Простейшие бактерии названия. Микробы. их имена. Что такое бактерии
Примеры бактерий и их особенности
Среды жизни бактерий: наземно-воздушная, водная, почвенная, организменная.
Характеризуются наличием прочной оболочки клеток, в состав надмембранного комплекса которой входит муреин (гликопротеин ).
Бактерии, оболочка которых богата муреином, окрашиваются анилиновыми красителями и йодом. Окрашивание не исчезает после обработки спиртом. Бактерии с невысоким содержанием муреина после окрашивания и обработки спиртом обесцвечиваются. Эти различия впервые обнаружил голландский микробиолог Христиан Грам в 1884 году. С тех пор используется метод «окрашивания по Граму», а бактерии на основании использования данного метода называют грамположительными или грамотрицательными.
Клеточная стенка многих бактерий окружена слизистой капсулой, выполняющей защитную функцию. У большинства палочковидных и спиралевидных бактерий имеются органоиды передвижения - жгутики, состоящие из одной нити - микротрубочки (жгутики эукариот состоят из 11 нитей).
В цитоплазме бактериальной клетки отсутствуют мембранные органоиды, но есть рибосомы. В центре клетки располагается двунитчатая молекула ДНК, замкнутая в кольцо. Бактерии - гаплоидные организмы.
Бактерии размножаются простым делением, которое может происходить с высокой скоростью (клетки могут делиться через каждые 20 минут). Изредка у бактерий может происходить явление, напоминающее половое размножение, заключающееся в обмене факторами наследственности в процессе конъюгации.
При неблагоприятных условиях бактерии образуют споры, которые имеют плотную оболочку, пропитанную смолоподобными веществами
Споры бактерий устойчивы к воздействию высоких и низких температур, ультрафиолетового излучения. Благодаря способности образовывать смо ры, бактерии могут переживать неблагоприятные условия среды в течение десятков лет (возбудители сибирской язвы - до 30 лет).
В природе бактерии играют огромную роль в качестве редуцентом в цепях питания, участвуют в круговороте веществ в природе, влияют на концентрацию веществ в земной коре, на почвообразование.
В жизни человека широко используются бактерии молочно-кислого брожения для получения кисло-молочных продуктов, клубеньковые бактерии и азотобактер - в качестве искусственных живых удобрений. Широко используются бактерии в пищевой, кожевенной промышленности, в сельском хозяйстве, в медицине и других отраслях.
Особого внимания заслуживают бактерии, вызывающие заболевания человека, домашних животных и культурных растений. Способность патогенных бактерий вызывать заболевание и гибель других организмов (растений, животных, человека) обусловлена 3 факторами: интенсивным размножением, способностью разрушать ткани и органы, способностью вырабатывать ядовитые вещества - токсины.
Бактерии изучаются специальной наукой - бактериологией. Вклад в развитие бактериологии внесли учёные: А. ван Левенгук, Л. Пастер, И. Листер, Н. А. Красильников, С. Н. Виноградский.
Сайт предоставляет справочную информацию исключительно для ознакомления. Диагностику и лечение заболеваний нужно проходить под наблюдением специалиста. У всех препаратов имеются противопоказания. Консультация специалиста обязательна!
Бактерии окружают нас всюду. Многие из них очень нужны и полезны человеку, а многие наоборот, вызывают страшные заболевания.Знаете ли Вы, каких форм бывают бактерии? А как они размножаются? А чем питаются? Хотите узнать?
.сайт) поможет Вам найти в этой статье.
Формы и размеры бактерий
Большинство бактерий – это одноклеточные организмы. Они отличаются большим разнообразием форм. В зависимости от формы бактериям даны и названия. Например, бактерии округлой формы называются кокками (всем известные стрептококки и стафилококки), бактерии в виде палочек называются бациллами, псевдомонадами или клостридиями (к бактериям такой формы относится знаменитая туберкулезная палочка или палочка Коха ). Могут бактерии иметь форму спиралек, тогда их имена спирохеты, вибриллы или спириллы . Не так часто, но случаются бактерии в форме звездочек, разных многоугольников или иных геометрических фигур.Бактерии совсем не велики, их размеры колеблются от половины до пяти микрометров. Самая большая бактерия имеет размер семьсот пятьдесят микрометров. После обнаружения нанобактерий оказалось, что их размеры намного меньше, чем ранее представляли себе ученые. Однако, на сегодняшний день, нанобактерии не слишком хорошо изучены. Некоторые ученые даже сомневаются в их существовании.
Агрегаты и многоклеточные организмы
Бактерии могут прикрепляться друг к другу при помощи слизи, образуя клеточные агрегаты. При этом каждая отдельная бактерия представляет собой самодостаточный организм, жизнедеятельность которого никак не зависит от приклеенных к ней сородичей. Иногда же бывает так, что бактерии приклеиваются для того, чтобы осуществить какую-то общую функцию. Некоторые же бактерии, как правило, нитчатой формы могут образовывать и многоклеточные организмы.Как они передвигаются?
Есть бактерии, которые сами не в состоянии передвигаться, но есть и такие, которые снабжены специальными устройствами для передвижения. Одни бактерии передвигаются при помощи жгутиков, а другие умеют скользить. Каким образом бактерии скользят, пока не до конца понятно. Есть мнение, что бактерии выделяют специальную слизь, которая облегчает скольжение. А еще есть бактерии, которые умеют «нырять». Для того чтобы опуститься в глубину какой-либо жидкой среды, такой микроорганизм может менять свою плотность. Чтобы бактерия начала движение в каком-то направлении, она должна получить раздражение.Питание
Есть бактерии, которые могут питаться лишь органическими соединениями, а есть такие, которые могут перерабатывать неорганику в органику и уже после этого использовать для собственных нужд. Энергию бактерии получают тремя способами: с использованием дыхания, брожения или фотосинтеза.Размножение
По поводу размножения бактерий можно сказать, что он тоже не отличается однородностью. Есть бактерии, которые не делятся на полы и размножаются простым делением или почкованием. Некоторые цианобактерии обладают способностью к множественному делению, то есть за один прием они могут выдать до тысячи «новорожденных» бактерий. Есть и бактерии, которые размножаются половым способом. Конечно же, у них все это происходит очень примитивно. Но при этом две бактерии передают новой клетке свои генетические данные – это главная особенность полового размножения.Бактерии, несомненно, заслуживают Вашего внимания не только потому, что вызывают множество болезней. Эти микроорганизмы были первыми живыми существами, которые населили нашу планету. История бактерий на Земле насчитывает почти четыре миллиарда лет! Самыми древними из существующих на сегодняшний день являются цианобактерии, они появились три с половиной миллиарда лет назад.
Испытать на себе полезные свойства бактерий Вы можете благодаря специалистам корпорации Тяньши, которые разработали для Вас
Бактерии — самая древняя группа организмов из ныне существующих на Земле. Первые бактерии появились, вероятно, более 3,5 млрд лет назад и на протяжении почти миллиарда лет были единственными живыми существами на нашей планете. Поскольку это были первые представители живой природы, их тело имело примитивное строение.
Со временем их строение усложнилось, но и поныне бактерии считаются наиболее примитивными одноклеточными организмами. Интересно, что некоторые бактерии и сейчас ещё сохранили примитивные черты своих древних предков. Это наблюдается у бактерий, обитающих в горячих серных источниках и бескислородных илах на дне водоёмов.
Большинство бактерий бесцветно. Только немногие окрашены в пурпурный или в зелёный цвет. Но колонии многих бактерий имеют яркую окраску, которая обусловливается выделением окрашенного вещества в окружающую среду или пигментированием клеток.
Первооткрывателем мира бактерий был Антоний Левенгук — голландский естествоиспытатель 17 века, впервые создавший совершенную лупу-микроскоп, увеличивающую предметы в 160-270 раз.
Бактерии относят к прокариотам и выделяют в отдельное царство — Бактерии.
Форма тела
Бактерии — многочисленные и разнообразные организмы. Они различаются по форме.
| Название бактерии | Форма бактерии | Изображение бактерии |
| Кокки | Шарообразная | |
| Бацилла |  | Палочковидная |
| Вибрион | Изогнутая в виде запятой | |
| Спирилла |  | Спиралевидная |
| Стрептококки |  | Цепочка из кокков |
| Стафилококки |  | Грозди кокков |
| Диплококки | Две круглые бактерии, заключённые в одной слизистой капсуле |
Способы передвижения
Среди бактерий есть подвижные и неподвижные формы. Подвижные передвигаются за счёт волнообразных сокращений или при помощи жгутиков (скрученные винтообразные нити), которые состоят из особого белка флагеллина. Жгутиков может быть один или несколько. Располагаются они у одних бактерий на одном конце клетки, у других — на двух или по всей поверхности.

Но движение присуще и многим иным бактериям, у которых жгутики отсутствуют. Так, бактерии, покрытые снаружи слизью, способны к скользящему движению.
У некоторых лишённых жгутиков водных и почвенных бактерий в цитоплазме имеются газовые вакуоли. В клетке может быть 40-60 вакуолей. Каждая из них заполнена газом (предположительно — азотом). Регулируя количество газа в вакуолях, водные бактерии могут погружаться в толщу воды или подниматься на её поверхность, а почвенные бактерии — передвигаться в капиллярах почвы.
Место обитания
В силу простоты организации и неприхотливости бактерии широко распространены в природе. Бактерии обнаружены везде: в капле даже самой чистой родниковой воды, в крупинках почвы, в воздухе, на скалах, в полярных снегах, песках пустынь, на дне океана, в добытой с огромной глубины нефти и даже в воде горячих источников с температурой около 80ºС. Обитают они на растениях, плодах, у различных животных и у человека в кишечнике, ротовой полости, на конечностях, на поверхности тела.
Бактерии — самые мелкие и самые многочисленные живые существа. Благодаря малым размерам они легко проникают в любые трещины, щели, поры. Очень выносливы и приспособлены к различным условиям существования. Переносят высушивание, сильные холода, нагревание до 90ºС, не теряя при этом жизнеспособность.
Практически нет места на Земле, где не встречались бы бактерии, но в разных количествах. Условия жизни бактерий разнообразны. Одним из них необходим кислород воздуха, другие в нём не нуждаются и способны жить в бескислородной среде.
В воздухе: бактерии поднимаются в верхние слои атмосферы до 30 км. и больше.
Особенно много их в почве. В 1 г. почвы могут содержаться сотни миллионов бактерий.
В воде: в поверхностных слоях воды открытых водоёмов. Полезные водные бактерии минерализуют органические остатки.
В живых организмах: болезнетворные бактерии попадают в организм из внешней среды, но лишь в благоприятных условиях вызываю заболевания. Симбиотические живут в органах пищеварения, помогая расщеплять и усваивать пищу, синтезируют витамины.
Внешнее строение
Клетка бактерии одета особой плотной оболочкой — клеточной стенкой, которая выполняет защитную и опорную функции, а также придаёт бактерии постоянную, характерную для неё форму. Клеточная стенка бактерии напоминает оболочку растительной клетки. Она проницаема: через неё питательные вещества свободно проходят в клетку, а продукты обмена веществ выходят в окружающую среду. Часто поверх клеточной стенки у бактерий вырабатывается дополнительный защитный слой слизи — капсула. Толщина капсулы может во много раз превышать диаметр самой клетки, но может быть и очень небольшой. Капсула — не обязательная часть клетки, она образуется в зависимости от условий, в которые попадают бактерии. Она предохраняет бактерию от высыхания.
На поверхности некоторых бактерий имеются длинные жгутики (один, два или много) или короткие тонкие ворсинки. Длина жгутиков может во много раз превышать разметы тела бактерии. С помощью жгутиков и ворсинок бактерии передвигаются.

Внутреннее строение
Внутри клетки бактерии находится густая неподвижная цитоплазма. Она имеет слоистое строение, вакуолей нет, поэтому различные белки (ферменты) и запасные питательные вещества размещаются в самом веществе цитоплазмы. Клетки бактерий не имеют ядра. В центральной части их клетки сконцентрировано вещество, несущее наследственную информации. Бактерии, — нуклеиновая кислота — ДНК. Но это вещество не оформлено в ядро.
Внутренняя организация бактериальной клетки сложна и имеет свои специфические особенности. Цитоплазма отделяется от клеточной стенки цитоплазматической мембраной. В цитоплазме различают основное вещество, или матрикс, рибосомы и небольшое количество мембранных структур, выполняющих самые различные функции (аналоги митохондрий, эндоплазматической сети, аппарата Гольджи). В цитоплазме клеток бактерий часто содержатся гранулы различной формы и размеров. Гранулы могут состоять из соединений, которые служат источником энергии и углерода. В бактериальной клетке встречаются и капельки жира.
В центральной части клетки локализовано ядерное вещество — ДНК, не отграниченная от цитоплазмы мембраной. Это аналог ядра — нуклеоид. Нуклеоид не обладает мембраной, ядрышком и набором хромосом.
Способы питания
У бактерий наблюдаются разные способы питания. Среди них есть автотрофы и гетеротрофы. Автотрофы — организмы, способные самостоятельно образовывать органические вещества для своего питания.
Растения нуждаются в азоте, но сами усваивают азот воздуха не могут. Некоторые бактерии соединяют содержащиеся в воздухе молекулы азота с другими молекулами, в результате чего получаются вещества, доступные для растений.
Эти бактерии поселяются в клетках молодых корней, что приводит к образованию на корнях утолщений, называемых клубеньками. Такие клубеньки образуются на корнях растений семейства бобовых и некоторых других растений.
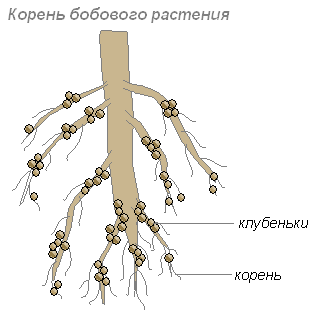
Корни дают бактериям углеводы, а бактерии корням — такие содержащие азот вещества, которые могут быть усвоены растением. Их сожительство взаимовыгодно.
Корни растений выделяют много органических веществ (сахара, аминокислоты и другие), которыми питаются бактерии. Поэтому в слое почвы, окружающем корни, поселяется особенно много бактерий. Эти бактерии превращают отмершие остатки растений в доступные для растения вещества. Этот слой почвы называют ризосферой.
Существует несколько гипотез о проникновении клубеньковых бактерий в ткани корня:
- через повреждения эпидермальной и коровой ткани;
- через корневые волоски;
- только через молодую клеточную оболочку;
- благодаря бактериям-спутникам, продуцирующим пектинолитические ферменты;
- благодаря стимуляции синтеза В-индолилуксусной кислоты из триптофана, всегда имеющегося в корневых выделениях растений.
Процесс внедрения клубеньковых бактерий в ткань корня состоит из двух фаз:
- инфицирование корневых волосков;
- процесс образования клубеньков.
В большинстве случаев внедрившаяся клетка, активно размножается, образует так называемые инфекционные нити и уже в виде таких нитей перемещается в ткани растения. Клубеньковые бактерии, вышедшие из инфекционной нити, продолжают размножаться в ткани хозяина.

Наполняющиеся быстро размножающимися клетками клубеньковых бактерий растительные клетки начинают усиленно делиться. Связь молодого клубенька с корнем бобового растения осуществляется благодаря сосудисто-волокнистым пучкам. В период функционирования клубеньки обычно плотные. К моменту проявления оптимальной активности клубеньки приобретают розовую окраску (благодаря пигменту легоглобину). Фиксировать азот способны лишь те бактерии, которые содержат легоглобин.
Бактерии клубеньков создают десятки и сотни килограммов азотных удобрений на гектаре почвы.
Обмен веществ
Бактерии отличаются друг от друга обменом веществ. У одних он идёт при участии кислорода, у других — без его участия.
Большинство бактерий питается готовыми органическими веществами. Лишь некоторые из них (сине-зелёные, или цианобактерии), способны создавать органические вещества из неорганических. Они сыграли важную роль в накоплении кислорода в атмосфере Земли.
Бактерии впитывают вещества извне, разрывают их молекулы на части, из этих частей собирают свою оболочку и пополняют своё содержимое (так они растут), а ненужные молекулы выбрасывают наружу. Оболочка и мембрана бактерии позволяет ей впитывать только нужные вещества.

Если бы оболочка и мембрана бактерии были полностью непроницаемыми, в клетку не попали бы никакие вещества. Если бы они были проницаемыми для всех веществ, содержимое клетки перемешалось бы со средой — раствором, в которой обитает бактерия. Для выживания бактерии необходима оболочка, которая нужные вещества пропускает, а ненужные — нет.
Бактерия поглощает находящиеся близ неё питательные вещества. Что происходит потом? Если она может самостоятельно передвигаться (двигая жгутик или выталкивая назад слизь), то она перемещается, пока не найдёт необходимые вещества.

Если она двигаться не может, то ждёт, пока диффузия (способность молекул одного вещества проникать в гущу молекул другого вещества) не принесёт к ней необходимые молекулы.
Бактерии в совокупности с другими группами микроорганизмов выполняют огромную химическую работу. Превращая различные соединения, они получают необходимую для их жизнедеятельности энергию и питательные вещества. Процессы обмена веществ, способы добывания энергии и потребности в материалах для построения веществ своего тела у бактерий разнообразны.
Другие бактерии все потребности в углероде, необходимом для синтеза органических веществ тела, удовлетворяют за счёт неорганических соединений. Они называются автотрофами. Автотрофные бактерии способны синтезировать органические вещества из неорганических. Среди них различают:
Хемосинтез
Использование лучистой энергии — важнейший, но не единственный путь создания органического вещества из углекислого газа и воды. Известны бактерии, которые в качестве источника энергии для такого синтеза используют не солнечный свет, а энергию химических связей, происходящих в клетках организмов при окислении некоторых неорганических соединений — сероводорода, серы, аммиака, водорода, азотной кислоты, закисных соединений железа и марганца. Образованное с использованием этой химической энергии органическое вещество они используют для построения клеток своего тела. Поэтому такой процесс называют хемосинтезом.
Важнейшую группу хемосинтезирующих микроорганизмов составляют нитрифицирующие бактерии. Эти бактерии живут в почве и осуществляют окисление аммиака, образовавшегося при гниении органических остатков, до азотной кислоты. Последняя, реагирует с минеральными соединениями почвы, превращаются в соли азотной кислоты. Этот процесс проходит в две фазы.
Железобактерии превращают закисное железо в окисное. Образованная гидроокись железа оседает и образует так называемую болотную железную руду.
Некоторые микроорганизмы существуют за счёт окисления молекулярного водорода, обеспечивая тем самым автотрофный способ питания.
Характерной особенностью водородных бактерий является способность переключаться на гетеротрофный образ жизни при обеспечении их органическими соединениями и отсутствии водорода.
Таким образом, хемоавтотрофы являются типичными автотрофами, так как самостоятельно синтезируют из неорганических веществ необходимые органические соединения, а не берут их в готовом виде от других организмов, как гетеротрофы. От фототрофных растений хемоавтотрофные бактерии отличаются полной независимостью от света как источника энергии.
Бактериальный фотосинтез
Некоторые пигментосодержащие серобактерии (пурпурные, зелёные), содержащие специфические пигменты — бактериохлорофиллы, способны поглощать солнечную энергию, с помощью которой сероводород в их организмах расщепляется и отдаёт атомы водорода для восстановления соответствующих соединений. Этот процесс имеет много общего с фотосинтезом и отличается только тем, что у пурпурных и зелёных бактерий донором водорода является сероводород (изредка — карбоновые кислоты), а у зелёных растений — вода. У тех и других отщепление и перенесение водорода осуществляется благодаря энергии поглощённых солнечных лучей.
Такой бактериальный фотосинтез, который происходит без выделения кислорода, называется фоторедукцией. Фоторедукция углекислого газа связана с перенесением водорода не от воды, а от сероводорода:
6СО 2 +12Н 2 S+hv → С6Н 12 О 6 +12S=6Н 2 О
Биологическое значение хемосинтеза и бактериального фотосинтеза в масштабах планеты относительно невелико. Только хемосинтезирующие бактерии играют существенную роль в процессе круговорота серы в природе. Поглощаясь зелёными растениями в форме солей серной кислоты, сера восстанавливается и входит в состав белковых молекул. Далее при разрушении отмерших растительных и животных остатков гнилостными бактериями сера выделяется в виде сероводорода, который окисляется серобактериями до свободной серы (или серной кислоты), образующий в почве доступные для растения сульфиты. Хемо- и фотоавтотрофные бактерии имеют существенное значение в круговороте азота и серы.
Спорообразование
Внутри бактериальной клетки образуются споры. В процессе спорообразования бактериальная клетка претерпевает ряд биохимических процессов. В ней уменьшается количество свободной воды, снижается ферментативная активность. Это обеспечивает устойчивость спор к неблагоприятным условиям внешней среды (высокой температуре, высокой концентрации солей, высушиванию и др.). Спорообразование свойственно только небольшой группе бактерий.

Споры — не обязательная стадия жизненного цикла бактерий. Спорообразование начинается лишь при недостатке питательных веществ или накоплении продуктов обмена. Бактерии в виде спор могут длительное время находиться в состоянии покоя. Споры бактерий выдерживают продолжительное кипячение и очень длительное проммораживание. При наступлении благоприятных условий спора прорастает и становится жизнеспособной. Спора бактерий — это приспособление к выживанию в неблагоприятных условиях.
Размножение
Размножаются бактерии делением одной клетки на две. Достигнув определённого размера, бактерия делится на две одинаковые бактерии. Затем каждая из них начинает питаться, растёт, делится и так далее.

После удлинения клетки постепенно образуется поперечная перегородка, а затем дочерние клетки расходятся; у многих бактерий в определённых условиях клетки после деления остаются связанными в характерные группы. При этом в зависимости от направления плоскости деления и числа делений возникают разные формы. Размножение почкованием встречается у бактерий как исключение.
При благоприятных условиях деление клеток у многих бактерий происходит через каждые 20-30 минут. При таком быстром размножении потомство одной бактерии за 5 суток способно образовать массу, которой можно заполнить все моря и океаны. Простой подсчёт показывает, что за сутки может образоваться 72 поколения (720 000 000 000 000 000 000 клеток). Если перевести в вес — 4720 тонн. Однако в природе этого не происходит, так как большинство бактерий быстро погибают под действием солнечного света, при высушивании, недостатке пищи, нагревании до 65-100ºС, в результате борьбы между видами и т.д.
Бактерия (1), поглотившая достаточно пищи, увеличивается в размерах (2) и начинает готовиться к размножению (делению клетки). Её ДНК (у бактерии молекула ДНК замкнута в кольцо) удваивается (бактерия производит копию этой молекулы). Обе молекулы ДНК (3,4) оказываются, прикреплены к стенке бактерии и при удлинении бактерии расходятся в стороны (5,6). Сначала делится нуклеотид, затем цитоплазма.

После расхождения двух молекул ДНК на бактерии появляется перетяжка, которая постепенно разделяет тело бактерии на две части, в каждой из которых есть молекула ДНК (7).
Бывает (у сенной палочки), две бактерии слипаются, и между ними образуется перемычка (1,2).

По перемычке ДНК из одной бактерии переправляется в другую (3). Оказавшись в одной бактерии, молекулы ДНК сплетаются, слипаются в некоторых местах (4), после чего обмениваются участками (5).
Роль бактерий в природе
Круговорот
Бактерии — важнейшее звено общего круговорота веществ в природе. Растения создают сложные органические вещества из углекислого газа, воды и минеральных солей почвы. Эти вещества возвращаются в почву с отмершими грибами, растениями и трупами животных. Бактерии разлагают сложные вещества на простые, которые снова используют растения.
Бактерии разрушают сложные органические вещества отмерших растений и трупов животных, выделения живых организмов и разные отбросы. Питаясь этими органическими веществами, сапрофитные бактерии гниения превращают их в перегной. Это своеобразные санитары нашей планеты. Таким образом, бактерии активно участвуют в круговороте веществ в природе.
Почвообразование
Поскольку бактерии распространены практически повсеместно и встречаются в огромном количестве, они во многом определяют различные процессы, происходящие в природе. Осенью опадают листья деревьев и кустарников, отмирают надземные побеги трав, опадают старые ветки, время от времени падают стволы старых деревьев. Всё это постепенно превращается в перегной. В 1 см 3 . поверхностного слоя лесной почвы содержатся сотни миллионов сапрофитных почвенных бактерий нескольких видов. Эти бактерии превращают перегной в различные минеральные вещества, которые могут быть поглощены из почвы корнями растений.
Некоторые почвенные бактерии способны поглощать азот из воздуха, используя его в процессах жизнедеятельности. Эти азотофиксирующие бактерии живут самостоятельно или поселяются в корнях бобовых растений. Проникнув в корни бобовых, эти бактерии вызывают разрастание клеток корней и образование на них клубеньков.
Эти бактерии выделяют азотные соединения, которые используют растения. От растений бактерии получают углеводы и минеральные соли. Таким образом, между бобовым растением и клубеньковыми бактериями существует тесная связь, полезная как одному, так и другому организму. Это явление носит название симбиоза.
Благодаря симбиозу с клубеньковыми бактериями бобовые растения обогащают почву азотом, способствуя повышению урожая.
Распространение в природе
Микроорганизмы распространены повсеместно. Исключение составляют лишь кратеры действующих вулканов и небольшие площадки в эпицентрах взорванных атомных бомб. Ни низкие температуры Антарктики, ни кипящие струи гейзеров, ни насыщенные растворы солей в соляных бассейнах, ни сильная инсоляция горных вершин, ни жёсткое облучение атомных реакторов не мешают существованию и развитию микрофлоры. Все живые существа постоянно взаимодействуют с микроорганизмами, являясь часто не только их хранилищами, но и распространителями. Микроорганизмы — аборигены нашей планеты, активно осваивающие самые невероятные природные субстраты.
Микрофлора почвы
Количество бактерий в почве чрезвычайно велико — сотни миллионов и миллиардов особей в 1 грамме. В почве их значительно больше, чем в воде и воздухе. Общее количество бактерий в почвах меняется. Количество бактерий зависит от типа почв, их состояния, глубины расположения слоёв.
На поверхности почвенных частиц микроорганизмы располагаются небольшими микроколониями (по 20-100 клеток в каждой). Часто они развиваются в толщах сгустков органического вещества, на живых и отмирающих корнях растений, в тонких капиллярах и внутри комочков.
Микрофлора почвы очень разнообразна. Здесь встречаются разные физиологические группы бактерий: бактерии гниения, нитрифицирующие, азотфиксирующие, серобактерии и др. среди них есть аэробы и анаэробы, споровые и не споровые формы. Микрофлора — один из факторов образования почв.
Областью развития микроорганизмов в почве является зона, примыкающая к корням живых растений. Её называют ризосферой, а совокупность микроорганизмов, содержащихся в ней, — ризосферной микрофлорой.
Микрофлора водоёмов
Вода — природная среда, где в большом количестве развиваются микроорганизмы. Основная масса их попадает в воду из почвы. Фактор, определяющий количество бактерий в воде, наличие в ней питательных веществ. Наиболее чистыми являются воды артезианских скважин и родниковые. Очень богаты бактериями открытые водоёмы, реки. Наибольшее количество бактерий находится в поверхностных слоях воды, ближе к берегу. При удалении от берега и увеличении глубины количество бактерий уменьшается.
Чистая вода содержит 100-200 бактерий в 1 мл., а загрязнённая — 100-300 тыс. и более. Много бактерий в донном иле, особенно в поверхностном слое, где бактерии образуют плёнку. В этой плёнке много серо- и железобактерий, которые окисляют сероводород до серной кислоты и тем самым предотвращают замор рыбы. В иле больше спороносных форм, в то время как в воде преобладают неспороносные.
По видовому составу микрофлора воды сходна с микрофлорой почвы, но встречаются и специфические формы. Разрушая различные отбросы, попавшие в воду, микроорганизмы постепенно осуществляют так называемое биологическое очищение воды.
Микрофлора воздуха
Микрофлора воздуха менее многочисленна, чем микрофлора почвы и воды. Бактерии поднимаются в воздух с пылью, некоторое время могут находиться там, а затем оседают на поверхность земли и гибнут от недостатка питания или под действием ультрафиолетовых лучей. Количество микроорганизмов в воздухе зависит от географической зоны, местности, времени года, загрязнённостью пылью и др. каждая пылинка является носителем микроорганизмов. Больше всего бактерий в воздухе над промышленными предприятиями. Воздух сельской местности чище. Наиболее чистый воздух над лесами, горами, снежными пространствами. Верхние слои воздуха содержат меньше микробов. В микрофлоре воздуха много пигментированных и спороносных бактерий, которые более устойчивы, чем другие, к ультрафиолетовым лучам.
Микрофлора организма человека
Тело человека, даже полностью здорового, всегда является носителем микрофлоры. При соприкосновении тела человека с воздухом и почвой на одежде и коже оседают разнообразные микроорганизмы, в том числе и патогенные (палочки столбняка, газовой гангрены и др.). Наиболее часто загрязняются открытые части человеческого тела. На руках обнаруживают кишечные палочки, стафилококки. В ротовой полости насчитывают свыше 100 видов микробов. Рот с его температурой, влажностью, питательными остатками — прекрасная среда для развития микроорганизмов.
Желудок имеет кислую реакцию, поэтому основная масса микроорганизмов в нём гибнет. Начиная с тонкого кишечника реакция становится щелочной, т.е. благоприятной для микробов. В толстых кишках микрофлора очень разнообразна. Каждый взрослый человек выделяет ежедневно с экскрементами около 18 млрд. бактерий, т.е. больше особей, чем людей на земном шаре.
Внутренние органы, не соединяющиеся с внешней средой (мозг, сердце, печень, мочевой пузырь и др.), обычно свободны от микробов. В эти органы микробы попадают только во время болезни.
Бактерии в круговороте веществ
Микроорганизмы вообще и бактерии в частности играют большую роль в биологически важных круговоротах веществ на Земле, осуществляя химические превращения, совершенно недоступные ни растениям, ни животным. Различные этапы круговорота элементов осуществляются организмами разного типа. Существование каждой отдельной группы организмов зависит от химического превращения элементов, осуществляемого другими группами.
Круговорот азота
Циклическое превращение азотистых соединений играет первостепенную роль в снабжении необходимыми формами азота различных по пищевым потребностям организмов биосферы. Свыше 90% общей фиксации азота обусловлено метаболической активностью определённых бактерий.
Круговорот углерода
Биологическое превращение органического углерода в углекислый газ, сопровождающееся восстановлением молекулярного кислорода, требует совместной метаболической активности разнообразных микроорганизмов. Многие аэробные бактерии осуществляют полное окисление органических веществ. В аэробных условиях органические соединения первоначально расщепляются путём сбраживания, а органические конечные продукты брожения окисляются далее в результате анаэробного дыхания, если имеются неорганические акцепторы водорода (нитрат, сульфат или СО 2).
Круговорот серы
Для живых организмов сера доступна в основном в форме растворимых сульфатов или восстановленных органических соединений серы.
Круговорот железа
В некоторых водоёмах с пресной водой содержатся в высоких концентрациях восстановленные соли железа. В таких местах развивается специфическая бактериальная микрофлора — железобактерии, окисляющие восстановленное железо. Они участвуют в образовании болотных железных руд и водных источников, богатых солями железа.
Бактерии являются самыми древними организмами, появившимися около 3,5 млрд. лет назад в архее. Около 2,5 млрд. лет они доминировали на Земле, формируя биосферу, участвовали в образовании кислородной атмосферы.
Бактерии являются одними из наиболее просто устроенных живых организмов (кроме вирусов). Полагают, что они - первые организмы, появившиеся на Земле.
Рис. 1. Человеческий организм на 90% состоит из микробных клеток. В нем содержится от 500 до 1000 всевозможных видов бактерий или триллионы этих удивительных жильцов, что составляет до 4-х кг совокупного веса.

Рис. 2. Бактерии, населяющие ротовую полость: Streptococcus mutants (зеленый цвет). Bakteroides gingivalis, вызывает периодонтит (сиреневый цвет). Candida albicus (желтый цвет). Вызывает кандидозы кожных покровов и внутренних органов.





Рис. 7. Микобактерии туберкулеза. Бактерии много тысячелетий вызывают заболевания у человека и животных. Туберкулезная палочка крайне устойчива во внешней среде. В 95% случаев передается воздушно-капельным путем. Чаще поражает легкие.

Рис. 8. Возбудитель дифтерии – коринебактерии или палочки Леффлера. Чаще развивается в эпителии слизистого слоя миндалин, реже гортани. Отек гортани и увеличенные лимфоузлы могут привести к асфиксии. Токсин возбудителя фиксируется на мембранах клеток сердечной мышцы, почек, надпочечников и нервных ганглиях и разрушает их.

Рис. 9. Возбудители стафилококковой инфекции. Патогенные стафилококки вызывают обширные поражения кожи и ее придатков, поражения многих внутренних органов, пищевую токсикоинфекцию, энтериты и колиты, сепсис и токсический шок.

Рис. 10. Менингококки – возбудители менингококковой инфекции. До 80% заболевших составляют дети. Инфекция передается воздушно-капельным путем от больных и здоровых носителей бактерий.

Рис. 11. Бордетеллы коклюша.

Рис. 12. Возбудители скарлатины стрептококки pyogenes.
Вредные бактерии микрофлоры воды
Местом обитания множества микробов является вода. В 1 см3 воды можно насчитать до 1 млн. микробных тел. Патогенные микроорганизмы попадают в воду от промышленных предприятий, населенных пунктов и животноводческих ферм. Вода с патогенными микробами может стать источником дизентерии, холеры, брюшного тифа туляремии, лептоспироза и др. Холерный вибрион и могут пребывать в воде достаточно много времени.
Рис. 13. Шигеллы. Возбудители вызывают бактериальную дизентерию. Шигеллы разрушают эпителий слизистой оболочки толстой кишки, вызывая тяжелый язвенный колит. Их токсины поражают миокард, нервную и сосудистую системы.

Рис. 14. . Вибрионы не разрушают клетки слизистого слоя тонкого кишечника, а находится на их поверхности. Выделяют токсин холероген, действие которого приводит к нарушению водно-солевого обмены в связи с чем организм теряет до 30 литров жидкости в сутки.

Рис. 15. Сальмонеллы — возбудители брюшного тифа и паратифов. Поражают эпителий и лимфоидные элементы тонкой кишки. С током крови попадают в костный мозг, селезенку и желчный пузырь, из которого вновь возбудители попадают в тонкий кишечник. В результате иммунного воспаления стенка тонкого кишечника разрывается и возникает перитонит.

Рис. 16. Возбудители туляремии (коккобактерии голубого цвета). Поражают респираторный отдел и кишечник. Обладают особенностью проникать в организм человека через целостные кожные покровы и слизистые глаз, носоглотки, гортани и кишечника. Особенность заболевания – поражение лимфоузлов (первичный бубон).

Рис. 17. Лептоспиры. Поражают капиллярную сеть человека, часто печень, почки и мышцы. Заболевание называют инфекционной желтухой.
Вредные бактерии микрофлоры почвы
Миллиарды «плохих» бактерий живет в почве. В 30-и сантиметровой толще 1-го гектара земли находится до 30-и тонн бактерий. Обладая мощным набором ферментов, занимаются расщеплением белков до аминокислот, тем самым принимают активное участие в процессах гниения. Однако эти бактерии приносят человеку немало неприятностей. Благодаря деятельности этих микробов очень быстро портятся продукты питания. Человек научился предохранять продукты длительного хранения путем стерилизации, засолки, копчения и замораживания. Некоторые виды этих бактерий способны испортить даже засоленные и замороженные продукты. попадают в почву от больных животных и человека. Некоторые виды бактерий и грибов пребывают в почве десятилетия. Этому способствует особенность этих микроорганизмов образовывать споры, которые долгие годы защищают их от неблагоприятных условий внешней среды. Они вызывают самые грозные заболевания – сибирскую язву, ботулизм, и столбняк.
Рис. 18. Возбудитель сибирской язвы. Десятилетия пребывает в почве в спорообразном состоянии. Особо опасная болезнь. Ее второе название – злокачественный карбункул. Прогноз заболевания неблагоприятный.

Рис. 19. Возбудитель ботулизма выделяет сильнейший токсин. 1 мкг этого яда убивает человека. Ботулотоксин поражает нервную систему, глазодвигательные нервы, вплоть до паралича и черепно-мозговые нервы. Смертность от ботулизма достигает 60%.

Рис. 20. Возбудители газовой гангрены очень быстро размножаются в мягких тканях организма без доступа воздуха, вызывая тяжелые поражения. В спорообразном состоянии сохраняется во внешней среде длительное время.

Рис. 21. Гнилостные бактерии.

Рис. 22. Поражение гнилостными бактериями продуктов питания.
Вредные бактерии, поражающие древесину
Ряд бактерий и грибов интенсивно разлагают клетчатку, играя важную санитарную роль. Однако среди них есть бактерии, вызывающие тяжелые заболевания животных. Плесневые грибы разрушают древесину. Деревоокрашивающие грибы окрашивают древесину в разные цвета. Домовой гриб приводит древесину в трухлое состояние. В результате жизнедеятельности этого гриба разрушаются деревянные постройки. Большой ущерб наносит деятельность этих грибов в разрушении животноводческих помещений.
Рис. 23. На фото видно, как домовой гриб разрушил деревянные балки перекрытия.

Рис. 24. Испорченный внешний вид бревен (синева), пораженных деревоокрашивающим грибом.

Рис. 25. Домовой гриб Merulius Lacrimans. а – ватообразная грибница; б – молодое плодовое тело; в – старое плодовое тело; г – старая грибница, шнуры и гниль древесины.
Вредные бактерии в пищевых продуктах
Продукты, обсемененные опасными бактериями, становятся источником кишечных заболеваний: брюшного тифа, сальмонеллеза, холеры, дизентерии и др. Токсины, которые выделяют стафилококки и палочки ботулизма , вызывают токсикоифекции. Сыры и все молочные продукты могут подвергнуться воздействию маслянокислых бактерий , которые вызывают маслянокислое брожение, в результате чего у продуктов появляется неприятный запах и цвет. Уксусные палочки вызывают уксусное брожение, что ведет к прокисанию вина и пива. Бактерии и микрококки, вызывающие гниение, содержат протеолитические ферменты, расщепляющие белки, чем придают продуктам дурно пахнущий запах и горький вкус. Плесенью покрываются продукты в результате поражения плесневыми грибами.
Рис. 26. Хлеб пораженный плесенью.

Рис. 27. Сыp пораженный плесенью и гнилостными бактериями.

Рис. 28. «Дикие дрожжи» Pichia pastoris. Фотография сделана с 600-кратным увеличением. Злостный вредитель пива. Повсеместно встречается в природе.
Вредные бактерии, разлагающие пищевые жиры
Маслянокислые микробы находятся повсюду. 25 их видов вызывают маслянокислое брожение. Жизнедеятельность жирорасщепляющих бактерий приводит к прогорканию масла. Под их воздействием прогоркают семена сои и подсолнечника. Маслянокислое брожение, которое вызывают эти микробы, портят силос, и он плохо поедается скотом. А влажное зерно и сено, пораженное маслянокислыми микробами, самосогревается. Влага, содержащаяся в сливочном масле, является хорошей средой, где размножаются гнилостные бактерии и дрожжевые грибы . Из-за этого масло портится не только снаружи, но и внутри. Если масло хранится долго, то на его поверхности могут поселиться плесневые грибы.
Рис. 29. Икорное масло, пораженное жирорасщепляющими бактериями.
Вредные бактерии, поражающие яйца и яичные продукты
В яйца бактерии и грибы проникают через поры наружной оболочки и ее повреждения. Наиболее чаще яйца инфицируются бактериями сальмонеллами и плесневыми грибами, яичный порошок — сальмонеллами и .
Рис. 30. Испорченные яйца.
Вредные бактерии в баночных консервах
для человека являются токсины ботулиновых палочек и палочек перфрингенс . Их споры проявляют высокую термоустойчивость, что позволяет микробам сохранять жизнедеятельность после пастеризации консервов. Находясь внутри банки, без доступа кислорода, они начинают размножаться. При этом выделяется углекислый газ и водород, от которых банка вздувается. Употребление в пищу такого продукта вызывает тяжелый пищевой токсикоз, который характеризуется крайне тяжелым течением и часто заканчивается смертью больного. Мясные и овощные консервы поражают уксуснокислые бактерии,Рис. 32. Во вздутых консервах могут находиться ботулиновые палочеки и палочки перфрингенс. Вздувает банку углекислый газ, который выделяют бактерии при размножении.
Вредные бактерии в зерновых продуктах и хлебе
Спорынья и другие плесневые грибы, которые поражают зерна, являются самыми опасными для человека. Токсины этих грибов термоустойчивы и не разрушаются при выпечке. Токсикозы, вызванные употреблением такой продукции, протекают тяжело. Мука, пораженная молочнокислыми бактериями , имеет неприятный вкус и специфический запах, комковатая на вид. Уже испеченный хлеб поражается бациллой субтилис (Вас. subtilis) или «тягучей болезнью». Бациллы выделяют ферменты, расщепляющие хлебный крахмал, что проявляется, вначале, не свойственным хлебу запахом, а потом липкостью и тягучестью хлебного мякиша. Зеленая, белая и головчатая плесень поражают уже испеченный хлеб. Распространяется при этом она по воздуху.
Рис. 33. На фото cпорынья пурпурная. Низкие дозы спорыньи вызывают сильные боли, умственные расстройства и агрессивное поведение. Высокие дозы спорыньи вызывают мучительную смерть. Ее действие связано с сокращением мышц под воздействием алкалоидов гриба.

Рис. 34. Грибница плесени.

Рис. 35. Споры зеленой, белой и головчатой плесени могут попасть из воздуха на уже испеченный хлеб и поразить его.
Вредные бактерии, поражающие фрукты, овощи и ягоды
Фрукты, овощи и ягоды обсеменяют почвенные бактерии, плесневые грибы и дрожжи, которые вызывают кишечные инфекции. Микотоксин патулин, который выделяют грибы рода Penicillium , способен вызывать раковые заболевания у человека. Yersinia enterocolitica вызывает заболевание иерсиниоз или псевдотуберкулез, при котором поражаются кожные покровы, желудочно-кишечный тракт и другие органы и системы.
Рис. 36. Поражение ягод плесневыми грибами.

Рис. 37. Поражение кожи при иерсиниозе.
Вредные бактерии проникают в организм человека с продуктами питания, через воздух, раны и слизистые оболочки. Тяжесть заболеваний, вызванных болезнетворными микробами, зависит от ядов, которые они вырабатывают и токсинов, возникающих при их массовой гибели. В течение тысячелетий они приобрели множество приспособлений, позволяющих им проникать и удерживаться в тканях живого организма и противостоять иммунитету.
Изучить вредное влияние микроорганизмов на организм и разработать профилактические мероприятия – вот задача человека!
БАКТЕРИИ (греческий bakterion палочка) - разнообразная по биол, свойствам группа широко распространенных на Земле микроскопических, в основном одноклеточных, организмов, принадлежащих к низшим формам жизни.
Первые сведения о бактериях были получены в 17 веке из исследований Левенгука, обнаружившего их основные формы. Бактерии могут существовать в самых разнообразных условиях.
Большинство их лишено хлорофилла. Исключение составляют анаэробные пурпурные и зеленые серобактерии, а также несерные пурпурные бактерии, содержащие хлорофилл и использующие солнечную энергию для фотосинтеза. Бактерии могут усваивать неорганический углерод и азот, использовать в качестве источников энергии многие неорганические и органические соединения, осуществлять превращения углерода, азота, серы, железа и других элементов.
Наряду с водорослями бактерии являются одними из самых древних организмов на Земле. Клеточное строение бактерий сходно с сине-зелеными водорослями, актиномицетами (см.) и спирохетами (см.), с которыми, как полагают, бактерии связаны филогенетически. Среди бактерий имеются виды, вызывающие заболевания у человека, животных и высших растений.
Систематика
Первые попытки классифицировать бактерии по морфологическим признакам были предприняты в 18 веке. Позднее в основу классификации были положены физиологические признаки. В качестве таксономических признаков применялись наиболее стабильные - форма, окраска по Tpainy (см. Грама метод), спорообразование, тип дыхания, биохимические, антигенные и другие свойства, однако до наст, времени не создано классификации, построенной на принципе филогенетического родства бактерий с учетом эволюционных связей.
Широкое распространение получила классификация Берджи (D. Bergey, 1957), в основу которой положены международные правила номенклатуры бактерий. Номенклатура выдержана в биноминальной системе, принятой в зоологических и ботанических классификациях (см. табл. 1). В качестве таксономических признаков взяты различные биологические свойства бактерий.
Таблица 1
|
Класс Schizomycetes |
||
|---|---|---|
|
семейство |
||
|
Патогенные бактерии |
||
|
Pseudomonadales (неподвижные клетки с полярными жгутиками) |
||
|
Eubacteriales (кокковидные, палочковидные бактерии с перитрихиальными жгутиками и неподвижные формы) |
||
|
Lactobacil laceae |
||
|
Peptostreptococcus |
||
|
Enterobacteriaceae |
||
|
Corynebacteriaceae |
||
|
Actinomycetales (нитевидные, ветвящиеся клетки - актиномицеты) |
Mycobacteriaceae |
|
|
Actinomycetaceae |
||
|
Streptomycetaceae |
||
|
Spirochaetales (подвижные, неригидные бактерии, у которых цитоплазма спирально закручена вокруг осевой нити) |
||
|
Mycoplasmatales (мелкие полиморфные, фильтрующиеся формы) |
Mycoplasmataceae |
|
|
Acholeplasmataceae |
||
|
Непатогенные бактерии |
||
|
Chlamydobacteriales |
||
|
Hyphomicrobiales |
||
Приведенные в таблице 1 микоплазмы - мельчайшие образования, отграниченные вместо ригидной клеточной стенки только цитоплазматической мембраной, существенно отличающиеся от бактерий, выделены в настоящее время в отдельный класс - Mollicutes (см. Mycoplasmataceae).
Морфология
Существует три основные формы бактерий - шаровидная, палочковидная и спиралевидная (рис. 1); большая группа нитчатых бактерий объединяет преимущественно водные бактерии и не содержит патогенных видов.
Шаровидные Ббактерии - кокки , подразделяются в зависимости от расположения клеток после деления на несколько групп: 1) диплококки (делятся в одной плоскости и располагаются парами); 2) стрептококки (делятся в одной плоскости, но при делении не отделяются друг от друга и образуют цепочки); 3) тетракокки (делятся в двух взаимно перпендикулярных плоскостях, образуя группы по четыре особи); 4) сарцины (делятся в трех взаимно перпендикулярных плоскостях, образуя группы кубической формы); 5) стафилококки (делятся в нескольких плоскостях без определенной системы, образуя скопления, напоминающие виноградные грозди). Средний размер кокков 0,5-1 мкм (см. Кокки).

Палочковидные бактерии имеют строго цилиндрическую или овоидную форму, концы палочек могут быть ровными, закругленными, заостренными. Палочки могут располагаться попарно в виде цепочек, но большинство видов располагается без определенной системы. Длина палочек варьирует от 1 до 8 мкм, средний диаметр 0,5-2 мкм. Принято собственно бактериями называть палочки, не образующие спор (см. Споры). Бактерии, образующие споры, называются бациллами. По принятой номенклатуре к бациллам относят аэробные формы. Анаэробных спорообразующих бактерий относят к клостридиям. Спорообразование у бацилл и клостридий не связано с процессом размножения. Споры у них относятся к типу эндоспор, представляющих собой круглые или овальные тела, преломляющие свет и окрашивающиеся по специальным методам (цветн. рис. 1 и 2). Расположение спор в клетке, их величина и форма характерны для каждого вида бактерий (рис. 2). Некоторые палочки (микобактерии, коринебактерии) образуют нитевидные особи, другие (клубеньковые бактерии) образуют разветвленные, звездчатые формы - так называемые бактероиды (рис. 3).
Спиралевидные формы бактерий подразделяют на вибрионы и спириллы. Изогнутость тел вибрионов не превышает одной четверти оборота спирали. Спириллы образуют изгибы из одного или нескольких оборотов (см. Вибрионы , Спириллы).
Некоторые бактерии обладают подвижностью, что отчетливо видно при наблюдении методом висячей капли (см.) или другими методами. Подвижные бактерии активно передвигаются с помощью особых органелл - жгутиков (см. Жгутики бактериальные) либо за счет скользящих движений (миксобактерии).

Капсула имеется у ряда бактерий и является их внешним структурным компонентом (рис. 4 и цветн. рис. 3). У ряда бактерий аналогично капсуле имеется образование в виде тонкого слизистого слоя на поверхности клетки. У некоторых бактерий капсула формируется в зависимости от условий их существования. Одни бактерии образуют капсулы только в макроорганизме, другие - как в организме, так и вне его, в частности на питательных средах, содержащих повышенные концентрации углеводов. Некоторые бактерии образуют капсулы независимо от условий существования (см. Капсульные бактерии). В состав капсулы большинства бактерий входят полимеризованные полисахариды, состоящие из пентоз и аминосахаров, уроновые кислоты, полипептиды и белки. Капсула не является аморфным образованием, а определенным образом структурирована. У некоторых бактерий, например, пневмококков, капсула определяет их вирулентность, а также некоторые антигенные свойства бактериальной клетки.
Клеточная стенка бактерий определяет их форму и обеспечивает сохранение внутреннего содержимого клетки. По особенностям химического состава и структуры клеточной стенки бактерии дифференцируют с помощью окрашивания по Граму.
Строение клеточной стенки различно у грамположительных и грамотрицательных бактерий. Основным слоем клеточной стенки, характерным для всех видов бактерий, является ригидный слой (синоним: мукопептидный слой, муреин, пептидогликан; последнее название наиболее соответствует химическому строению слоя), в состав которого вводят повторяющиеся остатки аминосахаров - N-ацетилглюкозамина и N-ацетилмурамовой кислоты, образующих основу линейного полимера - муреина.
С остатком N-ацетилмурамовой кислоты соединен полипептид, состоящий у большинства бактерий из четырех аминокислотных остатков - L-алани-на, D-глутаминовой кислоты, L-лизина или диаминопимелиновой кислоты (ДАП) и D-аланина в молярном отношении 1: 1: 1: 1. В составе пептида в зависимости от вида бактерий могут наблюдаться вариации. Лизин или ДАП могут быть заменены орнитином, 2,6-диаминобутаровой кислотой и др. Иногда к остатку глутаминовой кислоты присоединена добавочная аминокислота. Пептидные цепи соединены друг с другом с помощью перекрестных полипептидных цепочек, состав которых широко варьирует у разных видов бактерий. Перекрестные связи, например, у стафилококка, образованы пентаглициновыми мостиками, соединяющими D-аланин одной пептидной единицы с лизином другой. У некоторых бактерий перекрестные связи идентичны пептидным единицам. У E. coli пептидные цепи соединены непосредственно друг с другом через D-аланин одной цепи и ДАП другой. Схематическое изображение пептидогликана представлено на рис. 5.
У грамположительных бактериях в дополнение к пептидогликану имеются тейхоевые кислоты (рибит-тейхоевые и глицерин-тейхоевые), также образующие полимер и ковалентно связанные с пептидогликаном. У некоторых бактериях обнаружены тейхуроновые и 2-аминоманнуровая кислоты.
В состав клеточных стенок грамотрицательных бактерий, кроме ригидного слоя, входят липопротеиновый и липополисахаридный слои. Липополисахаридный слой (Л ПС) наиболее изучен у энтеробактерий, и особенно сальмонелл. Л ПС представляет собой комплекс фосфорилирование гетерополисахаридов, ковалентно связанных с содержащим глюкозамин липидом (липид А). В состав Л ПС входит О-антиген клетки (у энтеробактерий). Полисахаридная часть Л ПС состоит из основной (базисной) структуры и О-антигенной части. В состав базисной части, присущей всем энтеробактериям, входят гептоза, 2-кето-З-дезоксиоктонат (КДО), глюкоза, галактоза и N-ацетил-глюкозамин. Через КДО базисная часть присоединена к компоненту, состоящему из липида А, этанол амина, фосфата и КДО. С другой стороны (наружной) к базисной структуре присоединены боковые цепи, образованные повторяющимися олигосахаридными единицами. Наружные полисахаридные цепи видоспецифичны и являются соматическими О-антигенами. О-специфичность определяется углеводным составом всей боковой цепи, последовательностью расположения в ней углеводов и концевым сахаром, 6-дезокси- или 3,6-дидезоксигексозой. Наследственные нарушения в биосинтезе ЛПС энтеробактерий базисной части или O-боковых цепей приводят к появлению R-форм мутантов (см. Диссоциация бактерий).
Рис. 6. Строение клетки энтеробактерии (схематическое изображение): 1- детерминантные группы О-антигена; 2 - липопротеиновый слой; 3 - жгутик (Н-антиген); 4 - цитоплазматическая мембрана; 5 и б - рибосомы в цитоплазме; 7 - нуклеоид; 8-капсула; 9 - липополисахаридный слой; 10 - ригидный слой клеточной стенки.
Липопротеиновый слой (ЛП) у грамотрицательных бактерий, по представлению Вейделя (Weidel), является наружным слоем клеточной стенки. ЛПС занимает промежуточное положение, наиболее глубоко расположенным является ригидный слой. Эта схема не объясняет обнаружение О-антигена без предварительного разрушения Л П. Поэтому были предложены другие схемы строения стенки, согласно которым ЛП покрывает бактериальную клетку не сплошным слоем, а через него проходит ЛПС в виде «отростков», как это показано на рис. 6. Это представление подтверждено иммунохимическими методами с использованием ферритина при изучении локализации О-антигена.
У некоторых грамположительных бактериях клеточная стенка, так же как и у грамотрицательных, состоит не только из ригидного слоя, но имеет многослойное строение. Например, у стрептококков в ее состав входит белковый слой, промежуточный липополисахаридный и внутренний ригидный слой. Клеточная стенка не является инертной структурой в ферментативном отношении. В ее составе обнаружены аутолитические ферменты, фосфатаза, аденозинтрифосфатаза.
Цитоплазматическая мембрана бактерий прилегает к внутренней поверхности клеточной стенки, отделяет ее от цитоплазмы и является очень важным в функциональном отношении компонентом клетки. В мембране локализованы окислительно-восстановительные ферменты, с системой мембран связаны такие важнейшие функции клетки, как деление, биосинтез ряда компонентов, хемо- и фотосинтез и др. Толщина мембраны у большинства бактерий составляет 7-10 нм. Электронномикроскопическим методом обнаружено, что она состоит из трех слоев: двух электронно-плотных и промежуточного - электронно-прозрачного. В состав мембраны входят белки, фосфолипиды, липопротеины, небольшое количество углеводов и некоторых других соединений. Многие белки мембраны Б. являются ферментами, участвующими в процессах дыхания, а также в биосинтезе компонентов клеточной стенки и капсулы. В составе мембраны определяются также пермеазы, обеспечивающие перенос в клетку растворимых веществ. Мембрана служит осмотическим барьером, она обладает избирательной полупроницаемостью и ответственна за поступление внутрь клетки питательных веществ и выход из нее продуктов обмена.
Помимо цитоплазматической мембраны, в клетке бактерии имеется система внутренних мембран , получивших название мезосом, которые, вероятно, являются производными цитоплазматической мембраны; их строение варьирует у разных видов бактерий. Наиболее развиты мезосомы у грамположительных бактериях. Строение мезосом неоднотипно, их полиморфизм отмечается даже у одного и того же вида бактерий. Внутренние мембранные структуры могут быть представлены простыми инвагинациями цитоплазматической мембраны, образованиями в виде пузырьков или петель (чаще у грамотрицательных бактерий), в виде вакуолярных, ламмелярных, тубулярных образований. Мезосомы чаще всего локализованы у клеточной перегородки (рис. 7), отмечается также их связь с нуклеоидом. Поскольку в мезосомах обнаружены ферменты дыхания и окислительного фосфорилирования, многие авторы считают их аналогами митохондрий клеток высших. Предполагается, что мезосомы принимают участие в делении клетки, распределении дочерних хромосом в разделяющиеся клетки и спорообразовании. С мембранным аппаратом клетки связаны также функции фиксации азота, хемо- и фотосинтеза. Следовательно, можно полагать, что мембраны клетки играют определенного рода координирующую роль в пространственной организации ряда ферментных систем и органелл клетки.

Рис. 4 . Зерна волютина у коринебактерий

Цитоплазма и включения . Внутреннее содержимое клетки состоит из цитоплазмы (см.), представляющей собой сложную смесь различных органических соединений, находящихся в коллоидном состоянии. На ультратонких срезах цитоплазмы (рис. 7) обнаружено большое количество зерен, значительная часть которых является рибосомами. В цитоплазме бактерий могут содержаться внутриклеточные включения (цветн. рис. 4-6) в виде гранул гликогена, крахмала, жировых веществ. У ряда бактерий в цитоплазме находятся гранулы волютина, состоящие из неорганических полифосфатов, метафосфатов и соединений, близких к нуклеиновым кислотам. Роль волютина до конца не ясна. Некоторые авторы на основании его исчезновения при голодании клеток рассматривают волютин как запасные питательные вещества. Волютин обладает сродством к основным красителям, проявляет хромофильность и метахромазию, легко выявляется в клетках в виде крупных гранул, особенно при специальных методах окраски.
Рибосомы бактерии являются местом синтеза белков клетки, в процессе которого образуются структуры, состоящие из большого числа рибосом (до 20), называемые полирибосомами или чаще полисомами (рис. 8). В образовании полисом принимает участие м-РНК. По окончании синтеза данного белка полисомы вновь распадаются на одиночные рибосомы, или субъединицы. Рибосомы могут располагаться свободно в цитоплазме, но значительная их часть связана с мембранами клетки. На ультратонких срезах большинства бактерий рибосомы обнаруживаются в цитоплазме в виде гранул диаметром около 20 нм. Рибосомы E. coli, очищенные в присутствии ионов магния, осаждаются при ультрацентрифугировании со скоростью седиментации 70 S. При более низких концентрациях магния они диссоциируют на две субъединицы с константами седиментации 50 S и 30 S. Полагают, что 50 S частица имеет сферическую, а 30 S - уплощенную форму. При повышении концентрации ионов магния 70 S частицы образуют димеры. В свободном состоянии (вне синтеза белка) рибосомы находятся в диссоциированном состоянии в рибосомной фракции клеток. Диссоциацию рибосом на субъединицы стимулирует специальный фактор диссоциации. 50 S и 30 S субъединицы имеют мол. вес 1,8· 106 и 0,85-106 соответственно. Обе частицы состоят из рибосомальной РНК (или р-РНК) и белка. 50 S частица содержит по одной молекуле 23 S и 5 S р-РНК. 30 S частица содержит одну молекулу 16 S р-РНК. Белковый состав рибосом неоднороден. 30 S частицы состоят из двадцати одного, а 50 S из тридцати-тридцати пяти различных белков. Часть белков 30 S частиц рибосом нужна как для сборки рибосом, так и для их функционирования, другая часть важна только в функциональном отношении. Рибосомальная РНК имеет важное значение для правильной сборки и организации рибосом.
Степень агрегации рибосом регулируется ионами магния. В рибосомах найдены полиамины и рибонуклеаза I, участвующая, как полагают, в гидролизе м-РНК.
Рис. 10. Радиоавтография хромосомы бактерии coli. Видна циркулярнозамкнутая структура; слева вверху - схема репликации: X - начальная точка репликации, Y - точка роста; А - отреплецированный участок; Б - неотреплецированный участок; В - репликационная точка.
Ядро. Бактерии обладают дискретной ядерной структурой, в связи со своеобразием строения получившей название нуклеиида (рис. 9). Нуклеоиды Б. содержат основное количество ДНК клетки. Они окрашиваются методом Фейльгена (см. Дезоксирибонуклеиновые кислоты), хорошо видны при окраске по Романовскому - Гимзе (см. Романовского - Гимзы метод), после кислотного гидролиза или в живом состоянии при фазово-контрастной микроскопии, а также на ультратонких срезах в электронном микроскопе (рис. 7 и 9). Нуклеоид определяется в виде компактного одиночного или двойного образования. У растущих культур нуклеоиды часто выглядят в виде раздвоенных образований, что отражает их деление. Митотического деления ядерных структур у бактерий не обнаружено. Форма нуклеоидов и их распределение в клетке весьма изменчивы и зависят от ряда причин, в том числе и от возраста культуры. На электронных микрофотографиях в местах расположения нуклеоидов видны светлые участки меньшей оптической плотности. Ядерная вакуоль не отделена от цитоплазмы ядерной оболочкой. Форма вакуоли не постоянна. Ядерные участки заполнены пучками тонких нитей, образующих сложное переплетение. В составе ядерных структур бактерий не обнаружены гистоны (см.); предполагают, что их роль у бактерий выполняют полиамины. Ядра бактерий не похожи на ядра других организмов. Это послужило основой для выделения бактерий в группу прокариотов, в отличие от эукариотов, обладающих ядром, содержащим хромосомы, оболочку и делящимся путем митоза. Нуклеоид бактерии соединен с мезосомой. Характер связи пока не известен. Хромосома бактерий имеет циркулярно замкнутую структуру. Это было показано методом радиоавтографии у E. coli (рис. 10), предварительно меченных 3Н-тимидином. О структуре ДНК судили по распределению зерен меченого тимидина. Подсчитано, что длина замкнутой в кольцо ДНК клетки составляет 1100-1400 мкм, а молекулярный вес 2,8· 109 [Кернс (J. Cairns), 1963].
Жгутики и ворсинки . На поверхности некоторых бактерий имеются органеллы движения - жгутики (рис. 11). Их можно обнаружить с помощью особых методов окраски, микроскопирования в темном поле или в электронном микроскопе. Жгутики имеют спиралевидную форму, причем шаг спирали специфичен для каждого вида бактерий. На основании количества жгутиков и их расположения на поверхности клетки различают следующие группы подвижных микробов: монотрихи, амфитрихи, лофотрихи и перитрихи. Монотрихи имеют один жгутик, расположенный на одном из полюсов клетки и реже субполярно или латерально. У амфитрихов на каждом полюсе клетки расположено по одному жгутику. Лофотрихи имеют пучок жгутиков на одном или двух полюсах клетки. У перитрихов жгутики распределены без определенного порядка по всему телу клетки.
М. А. Пешков (1966) предлагает несколько другую терминологию. Амфи- и лофотрихи он объединяет термином «мультрихи» и выделяет смешанный тип, имеющий два или более жгутиков разного вида в разных точках прикрепления. Основание жгутиков (блефаропласт) расположено в цитоплазматической мембране. Жгутики почти целиком состоят из белка - флагеллина.
На поверхности некоторых бактерий (энтеробактерии), кроме жгутиков, имеются ворсинки (фимбрии, пили), видимые только под электронным микроскопом (рис. 12). Различают несколько морфологических типов ворсинок. Наиболее полно изучен первый тип (общий) и ворсинки, существующие только при наличии в клетке половых факторов (см. Половой фактор бактерий). Ворсинки общего типа покрывают всю поверхность клетки, состоят из белка; половых ворсинок приходится 1-4 на клетку. И те и другие обладают антигенной активностью (см. Конъюгация у бактерий).
Физиология
По химическому составу бактерии не отличаются от других организмов.

В состав бактерий входят углерод, азот, водород, кислород, фосфор, сера, кальций, калий, магний, натрий, хлор и железо. Их содержание зависит от вида бактерии и условий культивирования. Обязательным химическим компонентом клеток бактерий, как и других организмов, является вода, представляющая собой универсальную дисперсионную среду живой материи. Основная часть воды находится в свободном состоянии; ее содержание различно у разных бактерий и составляет 70-85% влажного веса бактерий. Кроме свободной, имеется ионная фракция воды и вода, связанная с коллоидными веществами. По составу органических компонентов клетки бактерий сходны с клетками других организмов, отличаясь, однако, наличием некоторых соединений. В состав бактерий входят белки, нуклеиновые кислоты, жиры, моно-, ди- и полисахариды, аминосахара и др. У бактерий имеются необычные аминокислоты: диаминопимелиновая (содержащаяся еще у сине-зеленых водорослей и риккетсий); N-метиллизин, входящий в состав флагеллина некоторых бактерий; D-изомеры некоторых аминокислот. Содержание нуклеиновых кислот зависит от условий культивирования, фаз роста, физиологического и функционального состояния клеток. Содержание ДНК в клетке более постоянно, нежели РНК. Нуклеотидный состав ДНК неизменен в процессе развития бактерий, видоспецифичен и используется как один из важнейших таксономических признаков. Бактериальные липиды разнообразны. Среди них встречаются жирные кислоты, фосфолипиды, воски, стероиды. Некоторые бактерии образуют пигменты (цветн. рис. 7-9) с интенсивностью, которая широко варьирует у одного и того же вида и зависит от условий выращивания. Твердые питательные среды более благоприятны для образования пигментов. По химическому строению различают каратиноидные, хиноновые, меланиновые и другие пигменты, которые могут быть красного, оранжевого, желтого, коричневого, черного, синего или зеленого цвета. Чаще пигменты нерастворимы в питательных средах и окрашивают только клетки. Пигменты, растворимые в воде (пиоцианин), диффундируют в среду, окрашивая ее. К пигментам бактерий относится также бактериохлорофилл, придающий фиолетовую или зеленую окраску некоторым фотосинтезирующим бактериям.
Ферменты бактерий делятся на функционирующие только внутри клетки (эндоферменты) и только вне клетки (экзоферменты). Эндоферменты в основном катализируют синтетические процессы, дыхание и т. п. Экзоферменты катализируют главным образом гидролиз высокомолекулярных субстратов до соединения с более низким молекулярным весом, способных проникать внутрь клетки.
В клетке ферменты связаны с соответствующими структурами и органеллами. Например, аутолитические ферменты связаны с клеточной стенкой, окислительно-восстановительные ферменты - с цитоплазматической мембраной, ферменты, связанные с репликацией ДНК,- с мембраной или нуклеоидом.
Активность ферментов зависит от ряда условий, в первую очередь от температуры выращивания бактерий и pH среды. Понижение температуры обратимо снижает, а повышение до определенных пределов (40-42°) повышает активность ферментов. У термофильных и психрофильных бактерий оптимум активности ферментов совпадает с оптимальной температурой роста. Оптимальная температура для мезофильных бактерий, к которым принадлежат патогенные бактерии, примерно равна 37°. Оптимум pH в основном лежит в пределах 4-7. Встречаются вариации оптимума pH. Ферменты бактерий, активность которых не зависит от присутствия субстрата в среде культивирования, называют конститутивными. Ферменты, синтез которых зависит от наличия субстрата в среде, называются индуцируемыми (старое название - адаптивные). Например, образование β-галактозидазы у кишечной палочки начинается только при добавлении в среду лактозы, которая индуцирует синтез этого фермента.
Контроль синтеза ферментов осуществляется путем ингибирования конечным продуктом или путем индукции и репрессии.
Ферментативная активность бактерий используется для их идентификации, чаще всего при этом изучаются сахаролитические и протеолитические свойства. Некоторые ферменты, образуемые патогенными бактериями, являются факторами вирулентности (см.).
Питание . Бактерии используют питательные вещества только в виде относительно небольших молекул, проникающих внутрь клетки. Такой способ питания, характерный для всех организмов растительного происхождения, называют голофитным. Сложные органические вещества (белок, полисахариды, клетчатка и др.) могут служить источником питания и энергии только после их предварительного гидролиза до более простых соединений, растворимых в воде либо в липоидах. Способность различных соединений проникать в цитоплазму клеток зависит от проницаемости цитоплазматической мембраны и химические структуры питательного вещества.
Вещества, которые служат источником питания бактерий, поразительно разнообразны. Важнейшим элементом, необходимым для живых организмов, является углерод. Одни виды бактерий (аутотрофы) могут использовать неорганический углерод из углекислоты и ее солей (см. Аутотрофные организмы), другие (гетеротрофы) - только из органических соединений (см. Гетеротрофные организмы). Подавляющее большинство бактерий относится к гетеротрофам. Для усвоения углерода требуется посторонний источник энергии. Немногочисленные виды бактерий, обладающие фотосинтетическими пигментами, используют энергию солнечного света. Эти бактерии называются фотосинтезирующими. Среди них имеются аутотрофы (зеленые и пурпурные серобактерии) и гетеротрофы (несерные пурпурные бактерии). Их называют также соответственно фотолитотрофами и фотоорганотрофами. Большинство же бактерий использует энергию химических реакций и называется хемосинтезирующими. Хемосинтезирующие аутотрофы называются хемолитотрофами, а гетеротрофы - хемоорганотрофами.
Гетеротрофные бактерии усваивают углерод из органических соединений различной химической природы. Легко усваиваются вещества, содержащие ненасыщенные связи или атомы углерода с частично окисленными валентностями. В связи с этим наиболее доступными источниками углерода являются сахара, многоатомные спирты и др. Некоторые гетеротрофы наряду с усвоением органического углерода могут усваивать и неорганический углерод.
Отношение к источникам азота также различно. Существуют бактерии, усваивающие минеральный и даже атмосферный азот. Другие бактерии неспособны синтезировать белковую молекулу или некоторые аминокислоты из простейших соединений азота. В этой группе имеются формы, использующие азот из отдельных аминокислот, из пептонов, сложных белковых веществ и из минеральных источников азота с добавлением несинтезируемых ими аминокислот. К этой группе принадлежат многие патогенные бактерии.
Дыхание . Часть веществ, проникающих внутрь бактериальной клетки, окисляясь, снабжает ее необходимой энергией. Этот процесс называют биол, окислением или дыханием.
Биологическое окисление сводится в основном к двум процессам: дегидрированию субстрата с последующим переносом электронов к конечному акцептору и накоплению в биологически доступной форме высвобождающейся энергии. Конечным акцептором электронов могут служить кислород, некоторые органические и неорганические соединения. При аэробном дыхании конечным акцептором электронов является кислород. Энергетические процессы, в которых конечным акцептором электронов является не кислород, а другие соединения, называются анаэробным дыханием, причем к собственно анаэробному дыханию некоторые исследователи относят те процессы, когда конечным акцептором электронов являются неорганические соединения (нитраты и сульфаты).
Под брожением понимают такие энергетические процессы, в которых органические соединения выступают одновременно как доноры и как акцепторы электронов.
Среди бактерий имеются строгие аэробы (см.), развивающиеся только в присутствии кислорода, облигатные анаэробы, развивающиеся только в отсутствие кислорода, и факультативные анаэробы (см.), способные к развитию и в аэробных и в анаэробных условиях. Большинство бактерий обладает пространственно организованной системой дыхательных ферментов, получившей название дыхательной цепи или цепи переноса электронов.
Дыхание у бактерий, подобно дыханию других организмов, сопряжено с процессами окислительного фосфорилирования, сопровождается образованием соединений, богатых макроэргическими связями (АТФ). Энергия, накапливающаяся в этих соединениях, используется по мере необходимости.
В качестве источника энергии бактерии могут использовать разнообразные органические соединения (углеводы, азотсодержащие вещества, жиры и жирные кислоты, органические кислоты и др.). Способность получать энергию в результате окисления неорганических соединений присуща лишь небольшой группе бактерий. Неорганические вещества, окисляемые ими, специфичны для каждого вида бактерий. К этим бактериям относятся нитрифицирующие бактерии, серобактерии, железобактерии и др. Среди них имеются и аэробы и анаэробы.
Фотосинтезирующие бактерии превращают энергию видимого света непосредственно в АТФ; этот процесс, осуществляемый в ходе фотосинтеза, называют фотофосфорилированием.
Рост и размножение
Бактериальная клетка начинает делиться после завершения последовательных реакций, связанных с воспроизведением ее компонентов.
Наиболее важным процессом роста клетки является воспроизведение ее наследственного аппарата. Разделению нуклеоида предшествуют процессы репликации ДНК (см. Репликация). Репликация начинается, когда отношение ДНК/белок клетки достигнет определенного уровня. Для инициации репликации требуется синтез специфических белковых продуктов. На реплицирующейся ДНК клетки при изучении ауторадиографическим методом различают две точки: точку начала репликации и точку роста (рис. 10). Репликативная точка продвигается по всей ДНК клетки, имеющей, как отмечалось, циркулярно замкнутую структуру. Время прохождения точки репликации от начала до конца всей циркулярной структуры ДНК, или время синтеза ДНК, постоянно и не зависит от скорости роста клеток. У быстро растущих культур, когда время генерации (время, протекающее между делением клеток) меньше, чем время, необходимое для репликации ДНК (40-47 минут у E. coli B/r), новая инициация начинается до окончания предыдущей. Таким образом, у быстро растущих культур имеется несколько репликационных точек (вилок). Процесс репликации ДНК сопровождается сегрегацией синтезирующихся цепей ДНК во вновь образующиеся дочерние клетки. В разделении нитей ДНК большую роль играют мезосомы клетки.
Рост палочковидных клеток во время цикла генерации сводится к экспоненциальному увеличению их длины. Во время деления рост клетки замедляется и начинается вновь после деления.
Окончание репликации ДНК является моментом, инициирующим деление клетки. Угнетение синтеза ДНК до окончания репликации приводит к нарушению процесса деления: клетка перестает делиться и растет в длину. На примере E. coli показано, что для начала деления требуется наличие термолабильного белка и такое соотношение между отдельными полиаминами в клетке, при котором количество путресцина должно превышать количество спермидина. Имеются данные о значении фосфолипидов и аутолизинов для процесса деления клеток.
У растущей культуры бактерий синтезируется полный набор рибосом. Рибосомальная РНК первоначально синтезируется на ДНК-матрице, затем модифицируется и превращается в зрелые 16 S и 23 S р-РНК. 5 S р-РНК также не является прямым продуктом транскрипции (см.). Предшественники рибосом не содержат полного набора рибосомальных белков. Полный набор появляется только в процессе созревания.
Механизм воспроизведения мезосом, как и мембранного аппарата клетки, еще не ясен. Предполагают, что при росте бактериальной клетки мезосомы постепенно разделяются.
При росте бактериальной клетки клеточная перегородка формируется рядом с мезосомой (рис. 7). Образование перегородки приводит к делению клетки. Вновь образованные дочерние клетки отделяются друг от друга. У некоторых бактерий образование перегородки не приводит к разделению клеток: образуются многокамерные клетки.
Получен ряд мутантов у E. coli, у которых клеточная перегородка образуется либо в необычном месте, либо наряду с перегородкой с обычной локализацией формируется добавочная перегородка близко от полюса клетки. В результате деления таких мутантов образуются как обычные клетки, так и маленькие клетки (мини-клетки) размером 0,3-0,5 мкм. Мини-клетки лишены, как правило, ДНК, так как при делении родительской клетки нуклеоид не попадает в них. В связи с отсутствием ДНК миниклетки используются в генетике бактерий для изучения выражения функции генов у внехромосомных факторов наследственности и других вопросов.
При выращивании в жидких питательных средах скорость роста популяции клеток изменяется во времени. Рост популяции бактерий разделяют на несколько фаз. После посева клеток в свежую питательную среду некоторое время бактерии не размножаются - эту фазу называют начальной стационарной или лаг-фазой. Лаг-фаза переходит в фазу положительного ускорения. В этой фазе начинается деление бактерий. Когда скорость роста клеток всей популяции достигает постоянной величины, начинается логарифмическая фаза размножения. В этот период можно рассчитать время генерации, количество генераций и некоторые другие показатели. Логарифмическая фаза сменяется фазой отрицательного ускорения, затем наступает стационарная фаза. Количество жизнеспособных клеток в данной фазе постоянно (М-концентрация - максимальная концентрация жизнеспособных клеток). Затем следует фаза отмирания популяции. На скорость роста популяции влияют: вид культуры бактерий, возраст посеянной культуры, состав питательной среды, температура выращивания, аэрация и др.
Во время роста популяции клеток в них накапливаются продукты обмена, происходит истощение питательных веществ и другие процессы, ведущие к переходу в стационарную и последующие фазы. При постоянном добавлении питательных веществ и одновременном удалении продуктов обмена можно добиться длительного пребывания клеток популяции в логарифмической фазе. Наиболее часто для этой цели применяют хемостат (см.).
Несмотря на постоянную скорость роста популяции бактерий в логарифмической фазе, отдельные клетки все же находятся в разных стадиях деления. Иногда важно синхронизировать рост всех клеток популяции, то есть получить синхронную культуру. Простыми методами синхронизации является изменение температурных условий или культивирование в условиях недостатка питательных веществ. Вначале культуру помещают в неоптимальные условия, затем сменяют их оптимальными. При этом у всех клеток популяции синхронизируется цикл деления, но синхронное деление клеток происходит обычно не более 3-4 циклов.
Ранее неоднократно выдвигались гипотезы, согласно которым превращение одних форм бактерий в другие в цикле развития идет по замкнутому кругу. Все эти гипотезы объединяются общим термином «циклогения». Теоретические представления о циклогении имеют в настоящее время лишь исторический интерес. Однако фактические данные о процессах диссоциации бактерий (см.) не потеряли своего значения.
Действие внешних факторов
Жизнеспособность бактерий при действии внешних факторов изучают разными методами, напр, путем подсчета выживших клеток. Для этого строят кривые выживаемости, выражающие зависимость числа выживших клеток от времени воздействия.
Бактерии относительно устойчивы к низким температурам. Бактерии более чувствительны к действию высоких температур. Обычно при прогревании бактерий при t° 60-70° происходит гибель вегетативных клеток, споры при этом не погибают. Чувствительность бактерий к высоким температурам используется при стерилизации (см.).
Разные виды бактерий относятся по-разному к высушиванию. Одни бактерии (например, гонококки) очень быстро погибают, другие (микобактерии) весьма устойчивы. Однако соблюдая определенные условия (наличие вакуума, специальных сред), можно получить высушенные лиофилизированные культуры бактерий, длительное время сохраняющие жизнеспособность (см. Лиофилизация).
Бактерии можно разрушить путем механического растирания с различными порошками (стеклянный, кварцевый), а также воздействием ультразвука.
Бактерии чувствительны к ультрафиолетовым лучам; наиболее эффективны лучи с длиной волны около 260 нм, что соответствует максимуму поглощения их нуклеиновыми кислотами. Ультрафиолетовые лучи обладают мутагенным действием. Рентгеновские лучи также обладают летальным и мутагенным действием (см. Мутагены).
Чувствительность к химиотерапевтическим препаратам и антибиотикам зависит от вида бактерий и механизма действия препарата на клетку. Из чувствительных бактерий могут быть получены устойчивые формы в результате мутации или при передаче факторов множественной лекарственной устойчивости микроорганизмов (см.).
Распространение бактерий в природе и их роль в круговороте веществ
Патогенность и вирулентность. Бактерии обитают в почве, воде, организме человека и животных. Разнообразные группы бактерий могут развиваться в условиях, не доступных для других организмов. Качественный и количественный состав бактерий, обитающих во внешней среде, зависит от многих условий: pH среды, температуры, наличия питательных веществ, влажности, аэрации, присутствия других микроорганизмов (см. Антагонизм микробов) и др. Чем больше в среде содержится разнообразных органических соединений, тем большее количество бактерий можно в ней обнаружить. В незагрязненных почвах и водах встречается сравнительно небольшое количество сапрофитных форм бактерий. В почве обитают спорообразующие и неспорообразующие бактерии, микобактерии, миксобактерии, кокковые формы. В воде встречаются разнообразные спорообразующие и неспорообразующие бактерии и специфические водные бактерии - водные вибрионы, нитчатые бактерии и др. В иле на дне водоемов обитают различные анаэробные бактерии. Среди бактерий, обитающих в воде и почве, имеются азотфиксирующие, нитрифицирующие, денитрифицирующие, расщепляющие целлюлозу бактерии. и др. В морях и океанах обитают бактерии, растущие при высоких концентрациях солей и повышенном давлении, встречаются светящиеся виды. В загрязненных водах и почве, кроме почвенных и водных сапрофитов, в большом количестве встречаются бактерии, обитающие в организме человека и животных,- энтеробактерии, клостридии и др.
Показателем фекального загрязнения обычно является наличие кишечной палочки. В связи с широким распространением бактерий и своеобразием метаболической активности многих их видов они имеют исключительно большое значение в круговороте веществ в природе. В круговороте азота участвуют многие виды бактерий - от видов, расщепляющих белковые продукты растительного и животного происхождения, до видов, образующих нитраты, которые усваиваются высшими растениями. Метаболическая активность бактерий обусловливает минерализацию органического углерода и образование углекислоты, возврат которой в атмосферу важен для поддержания жизни на Земле. Усвоение углекислоты из атмосферы производится зелеными растениями благодаря их фотосинтетической активности. Большая роль принадлежит бактериям в круговороте серы, фосфора, железа.
Сравнительно небольшая часть всех известных микробов способна вызывать заболевания человека и животных. Потенциальная способность бактерий вызывать инфекционные заболевания, являющаяся их видовым признаком, называется болезнетворностью или патогенностью. У одного и того же вида степень выраженности патогенных свойств может довольно широко варьировать. Степень патогенности штамма определенного вида бактерий называется его вирулентностью (см.). Среди бактерий имеются условно патогенные виды, болезнетворность которых зависит от состояния макроорганизма, внешней среды и др.
Генетика бактерий
Генетика бактерий - раздел общей генетики, изучающий наследственность и изменчивость у бактерий. Относительная простота организации бактерий, их способность расти в синтетических средах, быстрое размножение позволяют анализировать относительно редко возникающие изменения генома (см.) бактерий, составляющих многомиллиардные популяции, и проследить за их наследованием. Для этого используются специальные методы, обеспечивающие отбор из огромной популяции отдельных генетически измененных бактериальных клеток, передачу хромосомы или ее фрагментов от одних клеток (доноров) другим (реципиентам) с последующим генетическим анализом возникших рекомбинантов (см. Рекомбинация). Методы генетического анализа (см.) бактерий позволили изучить не только организацию бактериальной хромосомы, но и расшифровать тонкую структуру гена, а также установить функциональные взаимоотношения генетических единиц, составляющих отдельные бактериальные опероны (см.).
Развитие генетики бактерий связано с изучением бактериальной трансформации (см.), которое дало возможность установить роль ДНК как материальной основы наследственности. При изучении генетической трансформации у бактерий были разработаны методы экстракции и очистки ДНК, биохимические и биофизические методы анализа ее свойств. Это позволило не только изучать генетические изменения на клеточном уровне, но и сопоставлять эти изменения с изменением структуры ДНК. Таким образом, в совокупности с генетическими методами методы биохимического исследования генетического материала обеспечили возможность анализа закономерностей бактериальной генетики на молекулярном уровне.
Среди бактерий наиболее изученными в генетическом отношении являются кишечные палочки, у которых были открыты способы передачи генетического материала (хромосомы или ее фрагментов) от донора реципиенту, осуществляемые либо путем прямого скрещивания (см. Конъюгация у бактерий), либо с помощью бактериальных вирусов (см. Трансдукция). Другими микроорганизмами, обладающими такими же видами обмена генетическим материалом и по генетической характеристике приближающимися к кишечным палочкам, являются сальмонеллы.
Закономерности генетического обмена, установленные на кишечных палочках и сальмонеллах, присущи и ряду других микроорганизмов, играющих важную роль в инфекционной патологии. Феномены конъюгации и трансдукции обнаружены также у шигелл и некоторых других патогенных микроорганизмов, что позволяет осуществить генетический анализ факторов, обусловливающих их патогенность.
Для выяснения молекулярных механизмов, различных генетических феноменов значительный интерес представляют микроорганизмы, способные к генетической трансформации, при которой бактерии-реципиенты поглощают очищенную ДНК, экстрагированную из бактерий-доноров. В опытах трансформации выявляется генетическая активность изолированной, внеклеточной ДНК, что позволяет анализировать функциональную активность ДНК, подвергнутой различным воздействиям, изменяющим ее структуру как in vivo, так и in vitro.
Поэтому в молекулярно-генетических исследованиях широко используются трансформируемые виды бактерий, такие как Вас. subtilis, H. influenzae, Pneumococcus и др.
Свойства бактерий, как и любых других организмов, определяются набором присущих им генов. Запись генетической информации, закодированной в бактериальных генах, осуществлена на основе универсального триплетного кода (см. Генетический код). Яновским (С. Janofsky) были получены доказательства колинеарности (соответствия) между последовательностью нуклеотидов и последовательностью аминокислот в полипептиде и установлен in vivo состав отдельных триплетов, кодирующих включение различных аминокислот.
Набор генов, присущих бактериям, определяет их генотип (см.).Бактерии, обладающие одним и тем же генотипом, не всегда идентичны по своим свойствам; их свойства могут варьировать в зависимости от среды культивирования, возраста бактериальных культур, температуры выращивания и ряда других факторов внешней среды. Генотип определяет только потенциально присущие бактериальным клеткам свойства, выражение которых зависит от функционирования (активности) конкретных генетических структур. Хромосома бактерий включает 2 типа функционально различных генетических структур: структурные гены, детерминирующие специфичность белков, которые данная клетка способна синтезировать, и регуляторные гены, регулирующие активность структурных генов в зависимости от окружающих условий, в частности от наличия или отсутствия субстрата синтезируемого фермента либо от концентрации необходимого клетке соединения, от состояния генетического материала (репликация ДНК) и пр.
В активном состоянии структурные гены транскрибируются (см. Транскрипция), то есть становятся доступными для считывания генетической информации с помощью ДНК-зависимой РНК-полимеразы. Формирующаяся в процессе транскрипции информационная РНК (и-РНК) транслируется в соответствующий полипептид, структура которого закодирована в данных структурных генах.
По типу регуляции синтетические системы бактерий делят на 2 вида: катаболические и анаболические. Первые осуществляют утилизацию необходимой клетке энергии, вторые обеспечивают биосинтез соединений, необходимых бактериям.
Катаболическая система E. coli, осуществляющая расщепление лактозы на глюкозу и галактозу, детально изучена Жакобом и Моно (F. Jacob, J. Monod).
Ферменты этой системы (β-галактозидаза, галактозидпермеаза и галактозидтрансацетилаза) детерминируются соответствующими структурными генами. Рядом со структурными генами расположен регуляторный участок, так называемый оператор, «включающий» и «выключающий» считывание информации (транскрипцию) со структурных генов.
Другой регуляторной единицей данной системы является ген, контролирующий синтез репрессора - белка, способного соединяться с оператором. В присутствии репрессора структурные гены не транскрибируются РНК-полимеразой и синтеза соответствующих ферментов не происходит. Между оператором и геном-регулятором находится короткий участок ДНК - промотор - место посадки РНК-полимеразы. Добавленная к среде культивирования бактерий лактоза связывает репрессор, оператор оказывается свободным, и структурные гены начинают транскрибироваться, в результате чего происходит синтез ферментов. Таким образом, лактоза, являющаяся субстратом действия ферментов, выполняет функцию индуктора их синтеза.
Подобного рода регуляция свойственна и другим катаболическим системам. Синтез ферментов, индуцируемый субстратами их действия, называют индуцибельным.
Иного рода регуляция присуща анаболическим бактериальным системам. У этих систем ген-регулятор контролирует синтез неактивного репрессора-апорепрессора. При малых количествах конечного метаболита, контролируемого структурными генами данного биохимического пути (например, какой-нибудь аминокислоты), апорепрессор не соединяется с геном-оператором и не препятствует, следовательно, работе структурных генов и синтезу данной аминокислоты. В случае же избыточного образования конечного продукта последний начинает функционировать как корепрессор. Связываясь с апорепрессором, корепрессор превращает его в активный репрессор, соединяющийся с геном-оператором. В результате транскрибирование структурных генов и синтез соответствующих соединений прекращаются, то есть наблюдается репрессия системы. В процессе расхода клеткой избыточного конечного метаболита активный репрессор вновь превращается в апорепрессор, ген-оператор освобождается и структурные гены вновь приобретают активность, то есть происходит дерепрессия системы.
Таким образом, генетическим системам обоего рода - катаболическим (индуцибельным) и анаболическим (репрессибельным) - свойственна регуляция по типу обратной связи: накопление и расход конечного продукта регулирует его синтез анаболическими системами; в катаболических системах в роли регулятора выступает субстрат действия синтезируемых ферментов.
Сдвиги в ходе клеточных синтетических процессов, вследствие которых могут возникать ненаследуемые изменения свойств бактерий одного и того же генотипа, могут быть выражены в различной степени в зависимости от окружающих условий. Резко нарушенные условия существования могут приводить к выключению функции отдельных структурных генов или их гиперфункции, что в свою очередь может привести к значительным морфологическим изменениям, несбалансированному росту и, в конечном счете, к гибели клеток.
Комплекс свойств бактерий, выявляемых в данных условиях существования, называют фенотипом. Фенотип бактерий хотя и зависит от окружающей среды, но контролируется генотипом, так как характер и степень возможных для данной клетки фенотипических изменений определяется набором генов, то есть генотипом.
Как структурные, так и регуляторные гены бактерии локализуются в бактериальной хромосоме и в сумме образуют генетический аппарат бактерий. Помимо этого, бактерии могут нести внехромосомные генетические детерминанты - плазмиды (см.), которые, как правило, не являются жизненно необходимыми для клетки. Напротив, активация функций некоторых из них (например, бактериоцинов) губительна для бактериальных клеток, не несущих плазмид. Вместе с тем плазмидные элементы придают бактериям ряд свойств, представляющх большой интерес, с точки зрения инфекционной патологии. Так плазмидными детерминантами может быть обусловлена множественная устойчивость к лекарственным веществам (см. R-фактор), выработка альфа-гемолизина и других бактериальных токсинов.
Хромосома бактерий, как и клеток высших организмов, локализована в ядре.
В отличие от клеток высших организмов, бактериальное ядро лишено оболочки и именуется нуклеоидом. Количество нуклеоидов в бактериальных клетках варьирует в зависимости от фазы роста культуры: число нуклеоидов у кишечных палочек максимально в быстро размножающихся культурах, находящихся в логарифмической фазе роста. В стационарной фазе роста кишечные палочки содержат по одному нуклеоиду. Хромосома бактерий представляет собой молекулу ДНК, замкнутую в кольцо, с молекулярным весом порядка 1,5 - 2 X 109 дальтон.

Рис. 13. Схема последовательности передачи генетического материала при конъюгации E. coli, иллюстрирующая кольцевую структуру бактериальной хромосомы. Буквами обозначены различные гены. Правая стрелка - последовательность передачи генов (В,Г,Д,Е,А,Б) реципиенту донорным штаммом 1; левая стрелка - последовательность передачи генов (Д,Г, В,Б,А,Е) реципиенту донорным штаммом 2.
Кольцевое строение бактериальной хромосомы установлено тремя методами: ауторадиографическим, электронномикроскопическим и генетическим. В первом случае получены ауторадиограммы кольцевых структур бактериальной ДНК, во втором - электронномикроскопические изображения изолированной кольцевой ДНК, в третьем - установлены закономерности генетического обмена, объяснимые только кольцевым строением хромосомы. Это можно пояснить следующим условным примером. Допустим, что в процессе скрещивания бактерий (конъюгации) от одной бактерии к другой передаются гены, обозначенные буквами А, Б, В, Г, Д, Е. Один из использованных донорных штаммов Hfr (сокращение от английского выражения high frequency of recombination - высокая частота рекомбинации) имеет точку начала передачи хромосомы в области гена В. В этом случае наблюдается следующий порядок передачи генов: В, Г, Д, Е, А, Б. Второй штамм Hfr начинает передачу хромосомы с гена Д и передает ее в направлении, противоположном предыдущему. Гены в этом случае передаются в следующем порядке: Д, Г, В, Б, А, Е. Экспериментально показанное сохранение последовательности передачи генов при измененном порядке их переноса легко объясняется кольцевым строением хромосомы (рис. 13).
Методы, дающие возможность экспериментально осуществлять перенос генетического материала у бактерий (конъюгация, трансдукция и трансформация), позволили построить генетическую карту бактериальной хромосомы, отражающую относительную локализацию генов. С целью генетического картирования широко используется конъюгация, при которой реципиенту передаются большие участки бактериальной хромосомы, а иногда и вся хромосома донора. При конъюгационном картировании применяют различные подходы: устанавливают передачу отдельных генов по времени, выявляют сцепленную передачу генов, устанавливают частоту передачи генов, не подвергаемых селекции (неселективных), расположенных проксимально и дистально относительно селекционируемого гена, и т. п. Конъюгация, однако, в большинстве случаев не обеспечивает возможности достаточно точного картирования, так как при этом рекомбинация (см.) осуществляется на относительно протяженных участках хромосомы. Точное картирование производят с помощью трансдукции, при которой передаются более короткие фрагменты хромосомы бактерии, не превышающие 0,01 ее длины. Один из основных приемов трансдукционного картирования состоит в определении возможности котрансдукции (то есть совместной передачи) картируемого гена и гена, локализация которого в хромосоме известна. Наличие котрансдукции свидетельствует о близком (сцепленном) расположении анализируемых генов. С помощью трансдукции можно также устанавливать порядок расположения генов. Для этой цели используют специальный метод генетического анализа - так называемый тест трех точек, при котором анализ скрещиваний ведется относительно трех генов.
Трансформация для картирования используется сравнительно редко. Обработка реципиентных бактерий трансформирующей ДНК дает возможность передавать только очень небольшие участки бактериальной хромосомы. Вследствие этого с помощью трансформации можно анализировать лишь гены, которые составляют группы сцеплений.
Генетическая карта E. coli К-12, построенная на основе многолетних генетических исследований, проведенных в различных лабораториях мира, насчитывает в настоящее время несколько сот локализованных генов.
Рис. 14. Циркулярная генетическая карта, показывающая расположение генов в хромосоме E. coli. Гены обозначены символами, расшифрованными в табл. 3. Цифры на внутренних поверхностях окружностей обозначают единицы длины карты (время, в течение которого данный ген передается при конъюгации), выраженные в минутах (от 0 до 90 мин.).
На рис. 14 представлена генетическая карта E. coli, опубликованная в 1970 году Тейлором (A. L. Taylor) в журнале «Bacteriological Reviews» (США). Для удобства ориентации окружность генетической карты, схематически изображающая хромосому, разделена на отрезки - минуты, в сумме составляющие время, необходимое для передачи в процессе конъюгации всей хромосомы. Для кишечных палочек это время составляет около 90 минут. Размещенные по окружности символы обозначают соответствующие гены и расшифрованы в таблице 3, которая включает около 2000 бактериальных генов, функции которых в жизнедеятельности бактериальной клетки в значительной мере исследованы. Сведения о локализации генов на хромосоме бактерий позволяют решать конкретные задачи практической микробиологии. Они служат необходимой предпосылкой для изучения вирулентности и патогенности бактерий, их устойчивости к лекарственным препаратам, возможности создания аттенуированных штаммов и для других целей. В расположении генов кишечной палочки и сальмонелл существует выраженная гомология.
В ряде случаев гены (цистроны), контролирующие отдельные стадии синтеза конечного метаболита, расположены в одном участке бактериальной хромосомы. Последовательность расположения генов при этом соответствует последовательности использования детерминируемых ими промежуточных соединений в ходе синтеза конечного метаболита. В том же участке хромосомы, где расположены структурные гены, могут располагаться и регуляторные генетические единицы, составляющие в совокупности с соответствующими структурными генами оперон (см.). Примером таких оперонов являются группы генов, обеспечивающих синтез гистидина, триптофана и т. д.
В других случаях структурные и регуляторные гены одного и того же биохимического пути размещаются в различных областях бактериальной хромосомы, примером чего могут служить гены, контролирующие синтез метионина, расщепление арабинозы, синтеза пуринов и др.
Изучение генетического обмена у бактерий не ограничивается целью генетического картирования. Возможность такого обмена используется и при получении полезных для человека новых штаммов бактерий. В частности, рекомбинация между патогенными и непатогенными бактериями может быть использована для конструирования аттенуированных штаммов, то есть штаммов с ослабленной вирулентностью, пригодных для изготовления живых вакцин. Такие штаммы могут быть получены из патогенных бактерий (например, из дизентерийных бактерий) при замещении генетической области (или областей), определяющей их патогенность, соответствующими участками хромосомы непатогенных бактерий (например, кишечных палочек). Для создания аттенуированных штаммов необходимо не только обеспечить возможность генетического обмена, но и предварительно изучить генетические основы патогенности, вирулентности, иммуногенности и картировать определяющие их гены. Только при этом условии может быть осуществлено конструирование полноценных вакцинных штаммов, утративших только вирулентность, но сохранивших свойства, обеспечивающие иммуногенность.
Генетический обмен у бактерий осуществляется и в естественных условиях их обитания, следствием чего является рекомбинационная изменчивость бактерий, проявляющаяся в образовании атипичных форм. Это обстоятельство придает изучению рекомбинационного процесса практический интерес, так как механизм образования, патогенетическое и диагностическое значение атипичных форм - актуальнейшие вопросы инфекционной патологии.
Помимо фенотипической и рекомбинационной изменчивости, бактериям свойственна мутационная изменчивость, то есть изменчивость, обусловленная мутациями, представляющими собой структурные перестройки генов, полную или частичную их утрату (делеций), не связанные с рекомбинациями. Бактерии широко используются для изучения закономерностей мутационного процесса. Мутация (см.), то есть изменение генотипа,- явление, вызываемое действием мутагенных агентов. Они являются основой для проведения всех генетических исследований, так как изучение функции генов, их картирование и другие генетические задачи могут быть решены только с помощью соответствующих мутантов. Характер бактериальных мутантов, формирующихся под влиянием мутагенных агентов, не зависит от механизма действия мутагенов (см.). Создавшееся же на первом этапе развития бактериальной генетики представление об адекватности мутационной изменчивости бактерий используемым мутагенам, то есть о специфическом действии последних, оказалось ошибочным, так же как ошибочной оказалась концепция о спонтанном характере мутационного процесса. Указанное представление исходило из того, что при действии агентов, вызывающих гибель основной части бактериальной популяции, исследователи получали мутации, соответствующие примененному агенту. Так, например, действие сульфаниламидов сопровождалось выделением сульфамидорезистентных мутантов, действие фагов - выделением фагорезистентных мутантов и т. д. Работами Лурия (S. Luria), Дельбрюка (М. Delbruck), Ледерберга (J. Lederberg) и Ньюкомба (H. Newcombe) было показано, что формирование таких мутантов происходит до добавления губительно действующего агента, а последний играет лишь роль селекционирующего фактора. Мутационные изменения в бактериальных популяциях возникают во многих генах, но селекционирующие агенты отбирают лишь соответственные мутации. Так, например, в мутирующей популяции бактерий могут содержаться мутанты различного рода: ауксотрофы - неспособные синтезировать какие-либо необходимые клетке соединения; мутанты, утратившие или приобретшие способность ферментации отдельных углеводов; резистентные к антибиотикам и т. д. При высеве такой популяции на среду с антибиотиком немутировавшие особи, так же как особи, несущие мутации, не имеющие отношения к антибиотикоустойчивости, не дадут роста. Вырастут на такой среде только бактерии, имеющие мутации в гене, определяющем соответствующую резистентность. Это, однако, не означает, что происхождение антибиотикоустойчивых мутантов связано с воздействием селекционирующего агента. Причиной возникновения резистентных мутантов, как и мутантов, оставшихся невыявленными на среде с антибиотиком, являются мутационные события, совершившиеся до воздействия селекционирующего агента. В свою очередь это не означает, что селекционирующий агент не может обладать мутагенной активностью, но при наличии таковой он индуцирует мутации не только в генах, соответствующих механизму его действия, но и, как любой другой мутаген, в самых разнообразных генах, а селекционирует лишь соответственно измененные бактерии.
Несостоятельность концепции о спонтанном мутировании бактерий была опровергнута на том основании, что при испытании многочисленных химических соединений и физических агентов, возможно, действующих на обычно культивируемые популяции бакткрий, было установлено, что мутагенная активность свойственна чрезвычайно широкому кругу факторов, в том числе естественным метаболитам бактерий. Действие этих факторов не всегда поддается контролю, но объясняет причину возникновения так называемых спонтанных мутаций.
Согласно современной концепции, спонтанные мутации представляют собой явление того же порядка, что и экспериментально полученные мутации, именуемые индуцированными. Как те, так и другие, причинно обусловлены. Различия состоят лишь в том, что индуцируемые мутации возникают под влиянием специально примененных мутагенных агентов, агенты же, вызывающие спонтанные мутации, остаются невыясненными. Термин «спонтанные», таким образом, не отражает суть явления и применяется условно для обозначения мутаций, возникающих без специальных воздействий.
Мутации, обусловленные воздействием мутагенных агентов, возникают в результате изменения последовательности нуклеотидов ДНК, проявлением чего является утрата или изменение функции полипептида, кодируемого данным геном, или изменение свойств регуляторных единиц бактериального генома (оператор, промотор). По «протяженности» различают генные и хромосомные мутации. Первые затрагивают один ген, вторые распространяются более чем на один ген. Хромосомные мутации возникают в результате выпадения большого числа нуклеотидов (делеций). Генные мутации чаще являются точечными, то есть заключаются в замене, вставке или выпадении одной пары нуклеотидов ДНК. Различают простые и сложные замены азотистых оснований в ДНК - транзиции и трансверзии (см. Мутация).
Бактериям свойственны прямые и обратные мутации. Последние часто имеют супрессорный характер. Мутагенные действия на бактериальные клетки оказывают все известные мутагены. Наиболее часто применяемыми мутагенами в бактериологических генетических исследованиях являются ультрафиолетовые лучи, проникающая радиация, моно- и бифункциональные алкилирующие агенты, аналоги оснований и ряд других.
Исследования последних лет, осуществленные на бактериях, вскрыли наличие генетически детерминированных систем, обеспечивающих репарацию повреждений генетического материала (ДНК). Этими исследованиями начато новое направление в генетике и молекулярной биологии. Данные, полученные при изучении бактериальной репаративной активности, привели к пересмотру ряда представлений о механизмах действия мутагенных агентов, формировании, фиксации и фенотипическом выражении мутационных изменений.
Антигены бактерий
Антигены бактерий локализованы в жгутиках, капсуле, клеточной стенке, мембранах и других структурах клетки. Антигены бактерий- биологически активные компоненты клетки, определяющие ее иммуногенные, токсические и инвазивные свойства. Расшифровка химической структуры бактериальных антигенов, контроля их синтеза клеткой и локализации в ней, а также иммуногенной специфичности является теоретической основой для создания эффективных методов диагностики и специфической иммунопрофилактики бактериальных инфекций.
Распределение антигенов в бактериальной клетке изучают иммуноцитологическими методами - специфической капсульной реакцией по Томчику (J. Tomcsik), прямым и непрямым методом флюоресцирующих антител, способом антител, меченных ферритином, йодом, ртутью или ураном, с применением электронной микроскопии ультратонких срезов, а также с помощью выделения отдельных структур для их последующего иммунологического изучения. Для выделения антигенов из бактерий применяют механическое разрушение при помощи мелких стеклянных шариков, ультразвук, высокое давление, детергенты, лизоцим или бактериофаг. Растворимые антигенные комплексы извлекают из бактерий путем их обработки протеолитическими ферментами, горячей водой, трихлоруксусной кислотой, диэтил гликолем, фенолом, мочевиной, пиридином, этиловым эфиром и др. Растворимые антигены очищают градиентным ультра-центрифугированием с помощью колоночной хроматографии или препаративным электрофорезом. Высокоочищенные антигены получают из энтеробактерий, коклюшных микробов, стрептококков и др.
Среди антигенов бактерий различают типо-, видо-, группо- и родоспецифические, а также «неспецифические». Большинство типо- и группоспецифических антигенов локализовано в жгутиках, капсуле и клеточной стенке бактерий. Антигены мембран и внутриклеточных структур бактериальных клеток изучены недостаточно.
Жгутиковые антигены (Н-антигены) представляют собой белок (флагеллин) с молекулярным весом 20 000- 40 000, состоящий из альфа- и бета-полипептидных цепей. При аналитическом ультрацентрифугировании флагеллин образует один гомогенный пик с коэффициентом седиментации 1,5-1,68. При нагревании до t° 100° в сильно кислой или щелочной среде жгутиковые антигены инактивируются. Предполагают, что аминокислотный состав разных серотипов жгутиковых антигенов сальмонелл, эшерихий и других энтеробактерий различен и это определяет их типовую специфичность. На различии в специфичности жгутиковых антигенов построена классификация серотипов сальмонелл. Изолированные жгутики энтеробактерий, холерного вибриона и других бактерий реагируют как Н-антиген (см. Жгутики бактериальные), однако фракция жгутиков содержит всегда примесь О-антигена. Жгутики и флагеллин S- и R-форм Proteus mirabilis содержат общий и различающийся антигенные компоненты. Антигенная специфичность зависит от соединения и последовательности субъединиц флагеллина жгутиковой нити. Методом иммунодиффузии (см.) у Н-антигена выявляются два компонента. При помощи препаративных иммунохимических методов можно получить Н-антиген, очищенный от О-антигена. Очищенный Н-антиген не обладает протективной активностью в опытах на лабораторных животных. Растворимые жгутиковые антигены используют для приготовления эритроцитарных Н-диагностикумов.
Капсульные антигены (К-антигены) многих бактерий типоспецифичны и стимулируют специфический иммунитет (см.). Многие из капсульных антигенов являются полисахаридами или мукопептидами.
Капсульные антигены пневмококков - типоспецифические полисахариды, в изолированном виде обладают свойствами гаптенов (см. Гаптены) и обозначаются как растворимая специфическая субстанция (SSS). В капсуле возбудителя сибирской язвы имеются гаптен-пептид, а также антигены белково-полисахаридной природы, чувствительные к протеолитическим ферментам. Капсульный глутамиловый полипептид, обнаруженный у Вас. megaterium, обладает свойствами антигена, перекрестно реагируя с антигенами клеточной стенки этого же микроба. Капсульные антигены полисахаридной природы выявлены у микробов рода Acetobacter. Эти антигены перекрестно реагировали с антисыворотками к стрептококкам группы В и G, а также к пневмококкам 23-го типа. Перекрестная серол, реакция обусловлена наличием в антигенах общей детерминантной группы - L-рамнозы.
Установлены перекрестные реакции между капсульными полисахаридными антигенами менингококков группы А и Вас. pumilus, менингококками группы С и E. coli 016: NM, пневмококками типа III и E. coli К7 и др.
В капсуле (точнее микрокапсуле) энтеробактерий обнаружены полисахаридные антигены: Vi-антиген (см.) у S. typhi, S. paratyphi С, E. coli, E. ballerup, В(К)-антигены у эшерихий, К-антигены у клебсиелл. У нек-рых сальмонелл обнаружены капсульные антигены белковой природы, которые обладают протективными свойствами (S. typhimurium, S. adelaide, Citrobacter). Капсульные полисахаридные антигены К. pneumoniae обладают адъювантным эффектом (см. Адъюванты).
В клеточной стенке многих видов микробов выявлены типо-, группо-, видо- и родоспецифические антигены. Согласно схеме Краузе (R. М. Krause, 1963) клеточная стенка стрептококка содержит типоспецифические белковые антигены (М-субстанция) и группоспецифические антигены полисахаридной природы. М-антиген (насчитывается до 60 типов) является протективным антигеном; в частично очищенном виде предложен в качестве вакцины. Проведенная амер. учеными проверка вакцины, состоящей из частично очищенного М-антигена, показала, что препарат вызывал у части привитых детей ревматизм. По данным ряда авторов, М-антиген тесно связан с антигеном, перекрестно реагирующим с антигеном сердечной мышцы человека. Предполагают, что перекрестно реагирующий антиген и М-антиген являются разными детерминантами одной белковой молекулы. Обнаружено также, что между M-антигеном стрептококка первого типа группы А и системой HLA лимфоцитов человека имеется связь. Другим группоспецифическим антигеном клеточной стенки стрептококков является мукопептидный антиген, специфичность которого обусловлена N-ацетилглюко-замином (для стрептококков группы А) и N-ацетилгалактозамином (для стрептококков группы С). Группоспецифическим антигеном молочнокислых стрептококков является внутриклеточная тейхоевая кислота.
В клеточной стенке стафилококков сосредоточены видоспецифические антигены - белковый А-антиген в поверхностном слое стенки и тейхоевая кислота, которая в комплексе с мукопептидом составляет внутренний слой стенки. А-антиген - преципитиноген, обнаруженный у большинства штаммов Staphylococcus aureus, его мол. вес 13 200. Он обладает способностью вступать в неспецифическую реакцию с Fc-фрагментом иммуноглобулинов класса G сыворотки крови человека и некоторых животных. Тейхоевая кислота - специфический преципитиноген, состоящий из субъединиц полирибитолфосфата, к которому присоединены N-ацетил глюкоз амин (детерминантная группа) и D-аланин. Тейхоевая кислота обнаружена в клеточных стенках стрептококков, стафилококков, микрококков Вас. subtilis и молочнокислых бактерий. Установлено, что тейхоевая кислота, изолированная из стафилококков, обладает протективными свойствами. Из клеточных стенок Cl. botulinum типа А изолирован и очищен белковый термостабильный антиген, устойчивый к действию трипсина.
В клеточных стенках коринебактерий, нокардий, микобактерий и актиномицетов обнаружены видо- и родоспецифические антигены. В состав мукопептида клеточной стенки коринебактерий, нокардий и микобактерий входит арабиноза и галактоза, которые обусловливают перекрестную серологическую реактивность между штаммами этих групп. В клеточной стенке дифтерийного микроба выявлены два антигена: поверхностный типоспецифический белок и более глубоко расположенный группоспецифический термостабильный полисахарид. Сложный набор антигенов выявлен в клеточной стенке анаэробных коринебактерий методом радиоиммуноэлектрофореза. Главным компонентом клеточных стенок этих микробов оказался кислый полисахарид. Группоспецифические мукополисахаридные гаптены выявлены в клеточных стенках Вас. anthracis. Эти гаптены реагируют в реакции преципитации с аналогичными антигенами, изолированными из Вас. cereus. Типоспецифические антигены Вас. megaterium также локализованы в клеточной стенке.
О - антиген (эндотоксин) энтеробактерий локализован в промежуточном слое клеточной стенки и представляет собой комплексное соединение, состоящее из белка или пептида, полисахарида и липида. Липополисахарид (глюцидолипоидный комплекс), извлеченный смесью фенола с водой, имеет молекулярный вес 106-107, на 60-70% состоит из фосфорилированного полисахарида и на 20-40% - из липида (липид А жирных кислот). Молекулярный вес очищенного полисахарида равен 20 000-60 000. Полисахарид О-антигенов разных видов энтеробактерий построен по одному принципу и состоит из базисной структуры и боковых S-специфических цепей, являющихся детерминантными группами. Базисная структура (иначе R-липополисахарид) всех серотипов сальмонелл включает глюкозамин, 2-кето-З-дезоксиоктанат (КДО), L-глицеро-D-манно-гептозу, галактозу и глюкозу.
Известно 6 хемотипов R-липополисахаридов, выявленных у соответствующих R-мутантов (Ra, Rb, Rc, Rd1, Rd2 и Re), которые различаются по степени дефектности в химической структуре. В состав белковых цепей входят 6-дезокси- и особенно 3,6-дидезоксигексозы. S-специфические боковые цепи построены из повторяющихся олигосахаридов. O-факторы представляют собой часть или всю детерминантную группу О-антигена. Они классифицируются по схеме Кауферманна- Уайта с помощью перекрестных или гомологичных реакций агглютинации. Концевой сахар, обладающий наибольшим сродством с активным центром антитела, обозначают как иммунодоминантный сахар. О-фактор 2 (группа А) детерминируется иммунодоминантным сахаром паратозой, О-фактор 4 (группа В) - абеквозой, О-фактор 9 (группа D) - тивелозой и т. д. Иммунодоминантным сахаром Shigella dysenteriae является рамноза. Специфичность О-антигенного комплекса обеспечивается не только иммунодоминантным сахаром, но и последовательностью расположения сахаров в боковой цепи и характером хим. связей между отдельными сахарами. Первоначально в микробной клетке синтезируется базисная структура полисахарида, а затем боковые цепи. Липидная часть О-антигена (липид А) у всех энтеробактерий почти идентична. Липид А представляет собой длинную цепь жирных кислот, производных полифосфо-d-глюкозамина, он прочно связан с O-специфическим полисахаридом. При этом биосинтез молекулы полисахарида, а также всей молекулы О-антигена генетически детерминирован.
Изолированный О-антиген (липополисахарид) имеет ветвистую структуру, которая нарушается при обработке комплекса дезоксихолатом натрия; образуются так называемые гаптенные субъединицы, из которых, по-видимому, построен весь комплекс. Изолированные О-антигены токсичны, пирогенны, вызывают местный и общий феномен Швартцмана (см. Швартцмана феномен), некроз опухолевидной ткани, специфическую и неспецифическую резистентность, а также обладают иммуностимулирующей и иммунодепрессивной активностью. Предполагают, что токсическая активность О-антигенов обусловлена липидом А. Введение О-антигена животным сопровождается лейкопенией и тромбоцитопенией. О-антиген вызывает феномен толерантности, сопровождающийся заметным увеличением фагоцитарной активности. Помимо О-антигена, в клеточных стенках энтеробактерий обнаружены лабильные к нагреванию антигены, а также общие антигены.
В 1962 г. Кунин (С. Kunin) с соавторами впервые описали общий антиген энтеробактерий, который по специфичности отличается от О-антигена. Общий антиген, извлеченный из E. coli 014,- полисахарид, вызывает у кроликов продукцию специфических антител.
Липополисахарид, или липид А, вводимый животному вместе с общим антигеном, подавляет продукцию антител к общему антигену. Другой вид общего антигена, названный Горжинским и Бродхаге (О. О. Gorzynski, 1964; О. О. Brodhage, 1961, 1962) С-антигеном, обнаружен у E. coli и Sh. sonnei. У Sh. sonnei при помощи реакции гемагглютинации выявлен бактериальный агглютиноген (ВА), связанный с липополисахаридом. В 1969 году Энгельбрехт (E. Engelbrecht) сообщил еще об одном общем антигене у энтеробактерий - «алкогольфильном» факторе, который был получен из S. paratyphi А и В, S. bareilly. Предполагается, что «алкогольфильный» антиген - полисахарид. В клеточных стенках у холерного вибриона локализован специфический альфа-антиген, у возбудителя коклюша - белковый протективный антиген и гистаминсенсибилизирующий фактор, у чумного микроба - антиген, извлекаемый смесью фенол - вода, и следы фракции I.
Протективная активность изолированных клеточных стенок продемонстрирована в опытах со стафилококками, стрептококками, туляремийных микробом, возбудителем чумы, энтеробактериями, коклюшным микробом, микобактериями, холерным вибрионом, бруцеллами. Из клеточных стенок указанных микробов извлекают растворимые антигены, обладающие протективной активностью. Клеточные стенки многих грамположительных и грамотрицательных микробов вызывают у лабораторных животных образование гранул, дерматиты, гепатиты, хроническик кардиты и артриты. В опытах in vitro клеточные стенки стимулируют выделение лизосомальных энзимов, обладают цитотоксическим эффектом, ингибируют флюцитоз бактерий и рост клеток.
Таким образом, поверхностные структуры многих бактерий содержат типо-, группо-, видо- и родоспецифические антигены, а также общие антигены для разных видов микробов. Многие из перечисленных антигенов имеют важное значение в патогенезе заболеваний и формировании специфического иммунитета.
Антигены мембран и внутриклеточных структур. В мембранах бактерий сосредоточены специфические антигены. Так, антигены цитоплазматической мембраны Вас. megaterium отличаются по своей специфичности от антигенов клеточной стенки.
Изучение антигенного строения мембран Micrococcus lysodeicticus показало, что на поверхности цитоплазматической мембраны расположено 8 антигенов. Во фракции мембран E. coli 0111: К 4: Н12 и других энтеробактерий обнаружены О- и Н-антигены, а также неидентифицированные антигены. Установлено, что 0-антиген мембран идентичен О-антигену клеточных стенок. Н-антиген мембран идентичен Н-антигену изолированных жгутиков, так как базальная часть жгутика прикреплена или расположена на внутренней поверхности цитоплазматической мембраны. Поэтому Н-антигенная активность мембран обусловлена антигенной активностью базальной части жгутика. Белки, извлеченные из мембран микоплазм разных серол, групп, обладали специфической антигенной активностью. Из коклюшного микроба, разрушенного ультразвуком, выделена палочковидная структура с коэффициентом седиментации 22s, которая обладает протективными свойствами (223-антиген). Этот антиген, вероятно, локализован в мембранах. Описан новый класс бактериального антигена - липотейхоевая кислота, которая может быть изолирована из стрептококков, молочнокислых бактерий и некоторых бацилл. Липотейхоевая кислота локализуется на поверхности цитоплазматической мембраны и является группоспецифическим антигеном. Липотейхоевая кислота построена из 25-30 глицерофосфатных остатков и липидного компонента (гликолипида). Часть глицерофосфатных остатков замещена глюкозой и D-аланином. Мембранные антигены большинства патогенных бактерий изучены мало.
Цитоплазматическая фракция бактерий отличается определенным своеобразием: в ней наряду с компонентами цитоплазмы (рибосомы, гранулы, фрагменты эндоплазматического ретикулума, клеточный сок) присутствуют компоненты ядра (ДНК и, возможно, ядерные белки).
Поэтому, подвергая цитоплазматическую фракцию иммунол, анализу, иногда трудно сказать, за счет каких антигенов выявлена активность.
Так называемая суммарная фракция цитоплазмы энтеробактерий, коклюшного микроба, кокков и других бактерий обладает слабой антигенной активностью. В цитоплазме ряда бактерий обнаружены общие антигены: между штаммами рода Nocardia и Streptomyces, Nocardia и Мусоbacterum. Идентичные цитоплазменные антигены выявлены у микобактерий, актиномицетов и коринебактерий. Однако в цитоплазме чумного микроба обнаружены специфические антигены: фракция I, «мышиный» токсин, VW-антиген и антигенный комплекс, извлекаемый трихлоруксусной обработкой. Перечисленные антигены могут иметь важное значение в патогенезе инфекции. На модели чумного микроба показано, что антигенные комплексы, получаемые методом фенол- вода, и антигенный комплекс, извлекаемый трихлоруксусной кислотой, являются разными антигенами и, возможно, локализованы в разных структурах. Из ультразвукового лизата шигелл Зельтман (G. Seltman, 1975) изолировал антиген, двигающийся, к аноду (АТА), который оказался общим для многих энтеробактерий. Этот антиген белковой природы, вероятно, расположен внутри клетки.
Выявлены антигены в рибосомах: в течение 1960-1963 годы было установлено, что в рибосомах бактерий локализованы три типа антигенов, общие для многих бактерий (по-видимому, РНК), общие для ограниченного числа видов (белок) и специфические для каждого вида. В 1967-1975 годы показано, что рибосомальные фракции, полученные из энтеробактерий, листерий, микобактерий, коклюшных микробов, холерных вибрионов, стафилококков, обладают протективными свойствами в опытах на лабораторных животных. При этом доказано, что протективная активность рибосом не связана с примесью антигенов клеточной стенки. Из рибосомальной фракции холерного вибриона методом ионообменной хроматографии изолирован белок, который обладал специфическими протективными свойствами, а очищенные рибосомы не вызывали защиты у животных. Однако одни исследователи предполагают, что протективная активность рибосом связана с РНК, другие - с белком, а третьи считают, что к изолированным рибосомам «прикреплен» какой-то углевод, возможно, клеточной стенки, который обладает специфическими свойствами антигена. Механизм защитного действия «рибосомальных» вакцин не выяснен.
Исследованиями Риби (E. Ribi) с сотр. было продемонстрировано наличие в цитоплазме энтеробактерий низкомолекулярного полисахарида, который по антигенным свойствам и хим. составу близок к О-антигену клеточной стенки. Этот полисахарид описан как плазматический. Его антигенная активность проявляется лишь тогда, когда он соединен с О-антигеном. Однако такой комплекс не вызывает образования антител у кроликов. Плазматический полисахарид был обозначен как нативный гаптен, построенный из «линейных молекул» (частиц), молекулярный вес которых равен 163 000, диаметр - 1,6 нм, длина - 130 нм. Молекулы нативного гаптена, в отличие от О-антигена, не образуют мицеллярных структур. Высказано предположение, что нативный гаптен является предшественником О-антигена клеточной стенки.
Многими исследователями установлено, что бактериальная ДНК обладает антигенными свойствами. Препараты бактериальной ДНК реагируют как антигены с гомологичной и гетерологичными сыворотками. Показана перекрестная серол, реактивность между ДНК бактерий и ДНК клеток макроорганизма.
Некоторые исследователи считают, что ДНК и нуклеопротеиды бактерий стимулируют аутоиммунный процесс.
Таким образом, бактерии имеют сложную мозаику антигенов, которые распределены почти во всех структурах и органелл ах. Одни из этих антигенов более активны, другие - менее. Наиболее важен с практической точки зрения вопрос выявления и выделения протективных антигенов в очищенном виде с целью изготовления эффективных вакцин и диагностических препаратов.
Библиогр.: Анатомия бактерий, пер. с англ., под ред. Г. П. Калины, М., 1960; Иерусалимский Н.Д. Основы физиологии микробов, М., 1963, библиогр.; Метаболизм бактерий, пер. с англ., под ред. В.А.Шорина, М., 1963, библиогр.; Многотомное руководство по микробиологии, клинике и эпидемиологии инфекционных болезней, под ред. H. Н. Шукова-Вережникова, т. 1, с. 58 и др., М., 1962; Пешков М. А. Цитология бактерий, М.- JI., 1955, библиогр.; он же, Сравнительная цитология сине-зеленых водорослей, бактерий и актиномицетов, М., 1966; Роуз Э. Химическая микробиология, пер. с англ., М., 1971, библиогр.; Станиславский Е. С. Бактериальные структуры и их антигенность, М., 1971, библиогр.; Bergey’s manual of determinative bacteriology, ed. by R. E. Buchanan a. N. E. Gibbons, Baltimore, 1975, bibliogr.; Annual Review of Microbiology, v. 1-26. Stanford, 1957-1972; Bacteria, ed. by I. C. Gunsalus a. R. Y. Stani-er, v. 1-5, N. Y.-L., 1960-1964; Helms tetter C.E. Sequence of bacterial reproduction, Ann. Rev. Microbiol., v. 23, p. 223, 1969, bibliogr.; К a e m p-fer R. a. Meselson M. Studies of ri-bosomal subunit exchange, Cold Spr. Harb. Symp. quant. Biol., v. 34, p. 209, 1969; Korn E.D. Cell membranes, structure and synthesis, Ann. Rev. Biochem., v. 38, p. 263, 1969; N о m u r e M. Bacterial ribosome, Bact. Rev., v. 34, p. 49, 1970; О s-born M. J. Structure and biosynthesis of the bacterial cell wall, Ann. Rev. Biochem., v. 38, p. 501, 1969; Replication of DNA in microorganisms, Cold Spr. Harb. Symp. quant. Biol., v. 33, 1968; R y t e r A. Association of the nucleus and membrane of bacteria, Bact. Rev., v. 32, p. 39, 1969; T о p 1 e у W. W. a. Wilson G. S. Principies of bacteriology and immunity, v. 1 - 2, Baltimore, 1964.
Генетика Б. - Браун В. Генетика бактерий, пер. с англ., М., 1968, библиогр.; Жакоб Ф.иВольман Э. Пол и генетика бактерий, пер. с англ., М., 1962; Захаров И. А. и Квитко К. В. Генетика микроорганизмов, JI., 1967; Сборник методик по генетике микроорганизмов, под ред. Р. Клауса и У. Хейса, пер. с англ., М., 1970, библиогр.; С к а-вронскаяА.Г. Мутации у бактерий, М., 1967, библиогр.; T а у 1 о г А. Z. а. T г о t-t e r C. D. Linkage map of Escherichia coli strain K-12,Bact. Rev., v. 36, p. 504, 1972, bibliogr.; CurtissR. Bacterial conjugation, Ann.Rev. Microbiol.,v. 23, p. 69, 1969; Hartman P. E., Hartman Z.a. Stahl R. Classification and mapping of spontaneous and induced mutations in the histidine operon of Salmonella, Advanc. Genet., v. 16, p. 1, 1971, bibliogr.; Proceedings of the 12-th international congress of genetics, v. 3, Tokyo, 1968; Sanderson Κ. E. Genetics of the Enterobacteriaceae, Advanc. Genet., v. 16, p. 35, 1971, bibliogr.
Антигены бактерий - Адо А. Д. и Федосеева В. Н. Локализация в клетках Neisseria perflava и Klebsiella pneumoniae антигенов, общих (перекрестно реагирующих) с тканями бронхо-легочного аппарата человека, Бюлл. Эксперим, биол, и мед., т. 81, Кя 3, с. 349, 1976; Гольдфарб Д. М. и Замчук Л.А. Иммунология нуклеиновых кислот, М., 1968, библиогр.; М и х а й-л о в И. Ф. Флюоресцирующие антитела и методы их применения, М., 1968, библиогр.; Петросян Е. А. Комплексные антигены тифо-паратифозной группы бактерий, М., 1961, библиогр.; Станиславский Е. С. Бактериальные структуры и их антигенность, М., 1971, библиогр.; H e у m e r В., S p a n e 1 R. a. Haferkamp O. Biologische Aktivitat bakterieller Zellwande, Immun. u. Infekt., Bd 3, S. 232, 1975; Luederitz O. a. o. Isolation and chemical and immunological characterization of bacterial lipopoly-saccharides, в кн.: Microbial toxins, ed. by Т. C. Montie, v. 4, p. 145, N. Y., 1971, bibliogr.; Owen P. a. Salton M. Antigenic and enzymatic architecture of Micrococcus lysodeikticus membranes established by crossed immunoelectrophoresis, Proc. nat. Acad. Sci. (Wash.), v. 72, p. 1711, 1975; Robbins J. B. a. o. Gross-reacting bacterial antigens and immunity to disease caused by encapsulated bacteria, в кн.: Immun. syst. a. infect. Dis., ed. by E. Ne-ter a. F. Milgrom, p. 218, Basel a. o., 1975; Wicken A. J. a. Knox K. W. Lipoteichoic acids, a new class of bacterial antigen, Science, v. 187, p. 1161, 1975.
B. С. Левашев; А. Г. Скавронская (ген. с табл.); Д. М. Гольдфарб (бакт. табл.). E. С. Станиславский.